El universo de las partículas subatómicas es un universo discreto, cuántico; es un universo de probabilidades, de incertidumbres, un universo de leyes que definitivamente no encajan en nuestra ilusión de un universo mecanicista, determinístico y continuo.
Esta nueva realidad incomodó a físicos de la talla de Albert Einstein. Es famosa su frase: “Dios no juega a los dados”, críticando la interpretación probabilística de la mecánica cuántica, especialmente al enfoque de la Escuela de Copenhague, representada por físicos de la talla de Niels Bohr y Werner Heisenberg. Pero también es famosa la respuesta de Bohr: “Einstein, deje de decirle a Dios lo que debe hacer”.
El velo que cubría la nueva física lo levantó accidentalmente Max Planck en 1900, durante su investigación de la radiación que emitía un cuerpo negro al calentarse, un fenómeno que no podía explicarse con la física clásica.
Durante su investigación, y para lograr que su modelo coincidiera con los resultados experimentales, partió de una premisa revolucionaria para la época: La energía no se intercambia de forma continua, la energía viene en “cuantos” o paquetes discretos, la energía de un oscilador atómico está cuantizada, y es proporcional a su frecuencia:
E=h⋅f
En esta ecuación h corresponde a una nueva constante, la “Constante de Planck.
El descubrimiento de dicha constante y la aplicación de los principios que llevaron a su determinación abrió una caja de pandora para la ciencia. En 1905 Albert Einstein pudo explicar el efecto fotoeléctrico a partir de dichas premisas, en 1925 le permitió a Erwin Schrödinger formular su famosa función de onda Ψ , y en 1926 condujo a que Max Born propusiera que |Ψ|² no representaba una densidad de energía (como pensaba Schrödinger), sino la “densidad de probabilidad” de encontrar una partícula en un punto en un momento determinado.
Entender los fundamentos de la mecánica cuántica ha sido de vital importancia para poder describir adecuadamente los procesos que operan al interior de la maquinaria fotosintética. Abordar esta nueva física es entonces un “debe” para quien desee profundizar realmente en el tema.
El desarrollo matemático que encabeza este blog es una invitación a abordar esta nueva física. M intención al escribir este blog es presentar de una manera sencilla el desarrollo de la función, resaltando en colores los aportes que para llegar a la misma tuvieron la relatividad especial, la mecánica cuántica y la física clasica.
Orlando Rodríguez
3 de julio , 2025
LECTURA ABSOLUTAMENTE RECOMENDADA:
David Derbes. A Student´s Guide to the Schrödinger Equation
Fig. 1. Diagrama general del supercomplejo fotosintético en bacterias púrpura
Las bacterias púrpura fotosintéticas son un grupo de Proteobacterias fotosintetizadoras anaerobias, las cuales utilizan como donadores de electrones succinatos, lactatos y eventualmente H2 o H2S a bajas concentraciones. Su aparato fotosintético, al igual que en los demás grupos biológicos, se encuentra embebido en invaginaciones de la membrana interna celular, las cuales generan un volumen confinado de periplasma donde se genera un gradiente de iones H+ requerido para la síntesis posterior de ATP.
El aparato fotosintético comprende un conjunto de proteinas transmembrana con funciones fínamente acopladas, e incluye complejos antena (LHII y LHI) destinados a la captura de energía lumínica y transporte de la misma al centro de reacción RC; el centro de reacción RC propiamente dicho, destinado a recibir y transferir la energía desde pigmentos fotosintéticos especiales hasta quinonas aceptoras a lo largo de una cadena de transporte de electrones, (Electron Transport Chain, ETC), incluye complejos cytocromo BC1, responsable de la generación PMF por medio de receptores especiales de quinonas; y finalmente complejos moleculares ATP synthase, que utilizan la PMF generada en la etapa anterior para ensamblar ATP.
El conjunto de subunidades conforman un supercomplejo transportador y almacenador de energía en forma de ATP y NADPH, moléculas fundamentales tanto para la activación de las cadenas de síntesis biológica como en los procesos de crecimiento, reproducción celular y en general en todas las funciones bioquímicas y fisiológicas demandantes de energía.
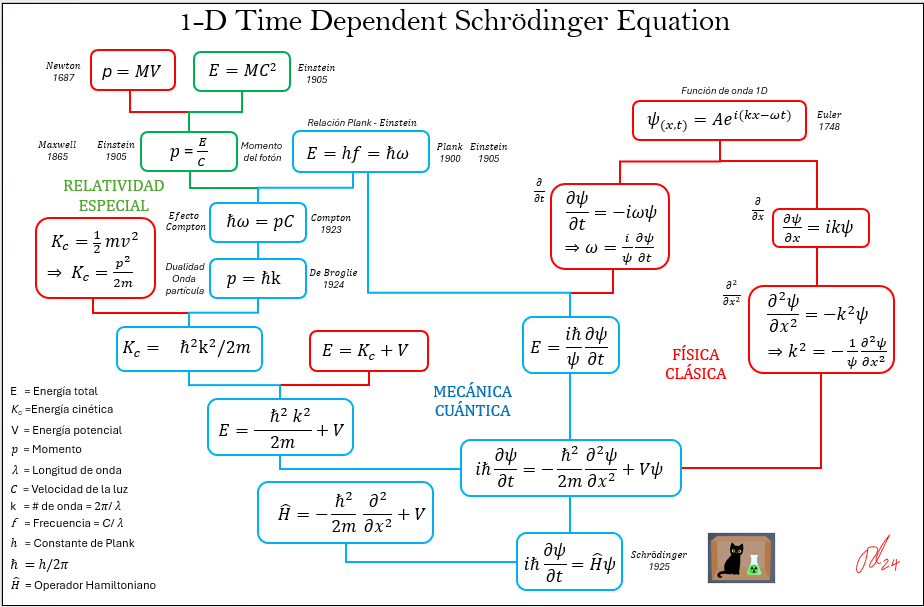
Fig. 2. Transferencia de energía entre complejos fotosintéticos. En la figura, “U” y “K” representan energías potencial y cinética, respectivamente.
CAPTURA DE ENERGÍA: COMPLEJOS ANTENA LHII Y LHI
La energía lumínica es capturada en arreglos concéntricos de carotenos y bacterioclorofilas (BChl) , localizados en complejos proteínicos octaméricos transmembrana, denominados complejos antena LHII (Light Harvest complex II). La energía es enrutada al interior de LHII de 500 a 800 a 850 nm para posteriormente exitar arreglos igualmente concéntricos de BChl ´s en LH1 a 875 nm.
Fig. 3. Light harvesting Complex II en Rhodoblastus acidophilus
Note en la figura la disposición en empalizada de los carotenos (moléculas café), y las disposiciones en anillos concéntricos de las 8 BChl´s paralelas a la superficie de la membrana plasmática (BChl´s violetas) y de las 16 perpendiculares a la misma (BChl´s azules).
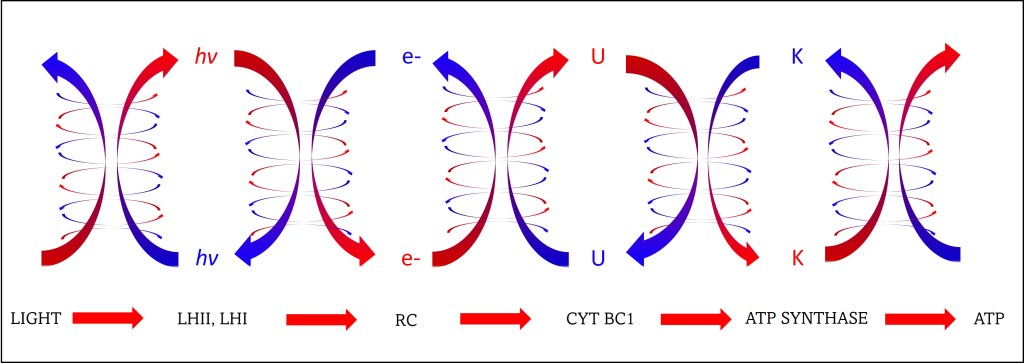
La transferencia de energía entre y dentro de anillos se realiza mediante “RESONANCIA DE FÖRSTER”, FRET, un tipo particular de resonancia cuántica no radiativa, que requiere distancias muy reducidas entre moléculas donadora y aceptora, y overlapping al menos parcial de la frecuencia de emisión del donador y frecuencia de exitación del aceptor, ver figuras 4 y 5.
Fig. 4. Overlapping parcial entre frecuencia de emisión del donador y exitación del aceptor
Fig. 5. Dependencia de la distancia entre moléculas donadora y aceptora para los valores de transferencia de energía.
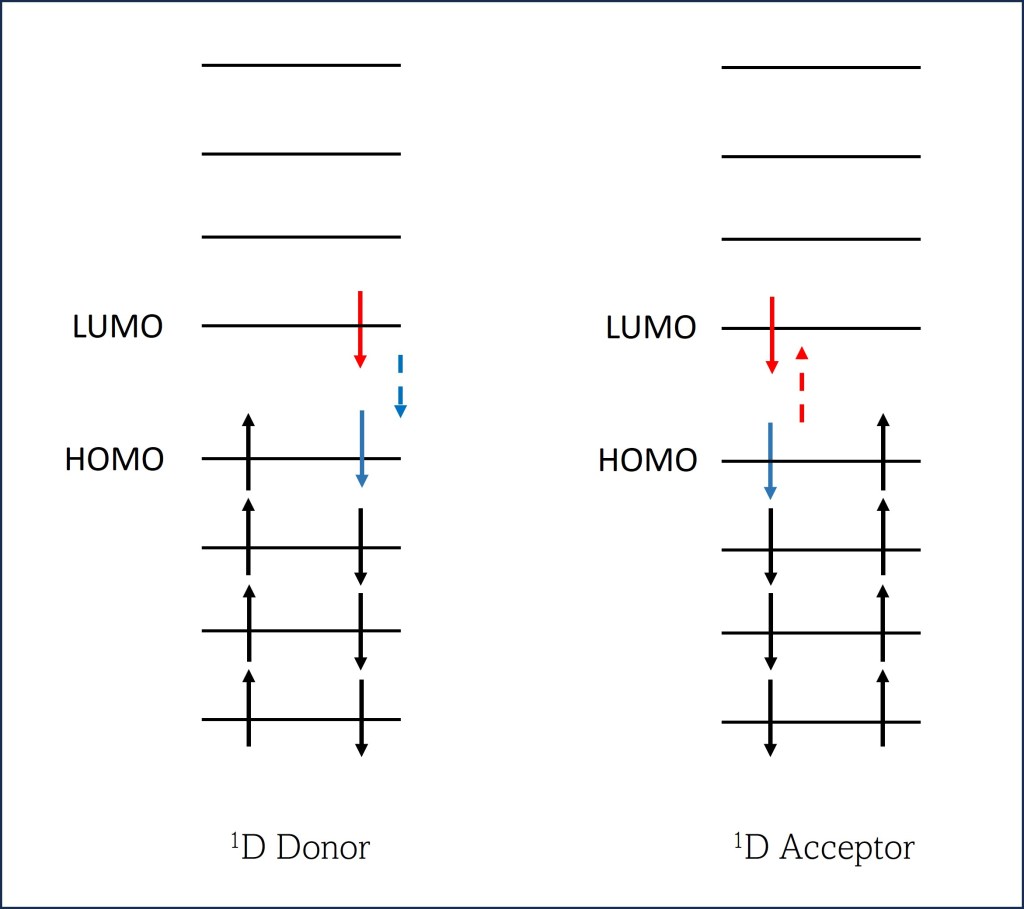
A diferencia de otros tipos de resonancia (Resonancia de Dexter), la Resonancia de Förster no mueve electrones exitados de donador a aceptor ni modifica su spin cuántico, por lo cual se define como una resonancia Singlet-Singlet no radiativa.
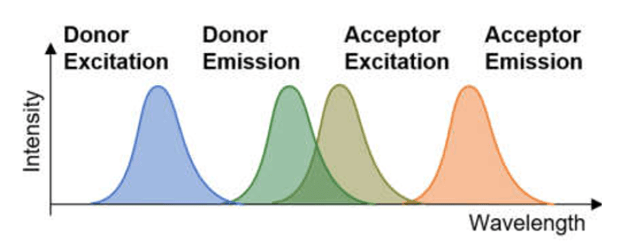
Partiendo de una exitación de un electrón de HOMO a LUMO en la molécula donadora, ver fig. 6., se transfiere posteriormente su energía a moléculas adyacentes mediante transferencia no radiativa.
Fig. 6. Exitación energética de HOMO a LUMO
En este caso, la molécula donadora retorna un electrón de LUMO (Lowest Unoccupied Molecular orbital) a Homo (Highest Occupied Molecular Orbital) mientra que en la molécula aceptora uno de sus electrones eleva su energía de HOMO a LUMO.
Fig. 7. Singlet-Singlet Förster Resonance Energy Transfer
Al menos dos propiedades cuánticas adicionales y probablemente una tercera son aprovechadas por los Complejos Antena para ejecutar su función: Superposición Cuántica, Coherencia Cuántica y eventualmente Superradiancia Cuántica.
Fig. 8.Transferencia de energía dentro de complejos (líneas azules) y entre complejos (Líneas rojas)
La Superposición Cuántica permite que multiples rutas probables coexistan tanto al interior de los complejos antena (flechas azules), como entre complejos (flechas rojas), garantizando que la energía sea transferida exitosamente y de manera rápida desde la molécula receptora del fotón hasta al Centro de Reacción RC sin que en el tránsito se “pierda”en rutas erráticas. Una vez alcanzado RC, la energía es “leída” por el par especial de clorofilas y en este momento la función de onda colapsa o elimina las demás rutas probables.
La unidireccional en cada paso también se garantiza por la disminución de frecuencias = Disminución de energía = aumento de longitud de onda durante el tránsito en el complejo de antenas (ver fig. 4).
Sin embargo también se debe notar que la exitación de LHI se dá a una mayor frecuencia (865 nm) que la frecuencia del donador (875 nm).
Es posible que la Coherencia y la Superradiancia Cuántica actuando en conjunto sean el elemento responsable del fenómeno.
Fig. 9.Superradiancia Cuántica
En el caso de los complejos antena, es probable que las moléculas individuales emisoras respondan bajo coherencia cuántica de forma espontánea , actuando cooperativamente como una única entidad. La coherencia lleva a que el grupo emita luz en un único haz coherente de alta intensidad. Este haz adquiere una intensidad N veces más fuerte que la intensidad esperada de un grupo de partículas independientes. En la figura 9 la variable N corresponde al número de partículas coherentes. Este efecto NO es basado en NINGUNA interacción entre las moléculas implicadas; es el resultado de las propiedades SIMÉTRICAS de la interacción del conjunto de partículas con el campo electromagnético lumínico global.
En resumen, los complejos antena, como parte del supercomplejo fotosintético, garantizan que la energía incidente de un fotón sea transmitida de manera eficiente hasta el centro de reacción RC, donde un par especial de clorofilas se exitarán al punto de liberar un electrón iniciando una cadena de transporte de electrones (ETC) que al final del proceso se acumulará en moléculas especializadas. El tránsito de fotónica a electrónica se ejecuta en el centro de reacción RC.
Orlando Rodríguez
Julio2, de 2025
LECTURAS RECOMENDADAS
Hu X, Schulten K. Model for the light-harvesting complex I (B875) of Rhodobacter sphaeroides. Biophys J. 1998 Aug;75(2):683-94. doi: 10.1016/S0006-3495(98)77558-7. PMID: 9675170; PMCID: PMC1299743.
Hu X, Ritz T, Damjanović A, Autenrieth F, Schulten K. Photosynthetic apparatus of purple bacteria. Q Rev Biophys. 2002 Feb;35(1):1-62. doi: 10.1017/s0033583501003754. PMID: 11997980.
Miroslav Z. Papiz, Anna M. Hawthornthwaite-Lawless, Steve M. Prince, Gerry McDermott, Andy A. Freer, Neil W. Isaacs, Richard J. Cogdell, A model for the photosynthetic apparatus of purple bacteria, Trends in Plant Science, Volume 1, Issue 6, 1996, Pages 198-206, ISSN 1360-1385
La fotosíntesis, como proceso bioquímico, tiene como función principal la captura de energía lumínica y el depósito final de la misma en moléculas orgánicas requeridas para soportar la estructura, mantenimiento y dinámica de la vida.
Siguiendo una estricta secuencia temporal, el proceso comprende las siguientes etapas:
Captura de energía en complejos especializados, denominados complejos antena, o Light Harvesting Complexes, LHC
Exitación electrónica en un par especial de clorofilas, en centros de reacción RC
Transporte de energía a lo largo de una cadena de reacciones de oxidoreducción, Electron Transport Chain (ETC)
Generación de un gradiente protónico (Protomotriz Force, PMF), en particular en el complejo cytocromo BC1
Utilización de PMF en ATP Synthase, para producción de ATP
Utilización de ATP en procesos de biosíntesis
A pesar de ser un proceso altamente eficiente, parte de la energía en cada etapa se pierde, bien en forma de calor, bien en procesos de disipación energética no fotoquímica no aprovechables (NPQ), de modo que el neto en cada etapa corresponde a la energía libre de Gibss para dicha etapa.
La evolución energética se ilustra en la gráfica a manera de rampas continuas, ascendentes o descendentes dependiendo del caso, sugiriendo funciones continuas (salvo el caso resaltado como línea punteada escalonada en ETC), sin embargo el proceso es discreto, con una “granularidad” dependiente del número de reacciones bioquimicas involucradas en cada etapa.
Aunque las arqueas se separaron evolutivamente de las bacterias hace al menos 3,000 – 3,200 millones de años, las bases de sus mecanismos de captura y transporte de energía se han conservado invariantes.
Así, el acoplamiento quimiosmótico entre un aparato generador de fuerza protomotiz (PMF) mediante bombeo de protones del interior hacia el exterior de la célula, y un complejo proteico (ATP-Synthase), diseñado específicamente para aprovechar este tipo de energía y acumularla en forma de ATP , son el común denominador para ambos grupos biológicos, si bien difieren de manera absoluta en la estructura y operación del complejo generador PMF.
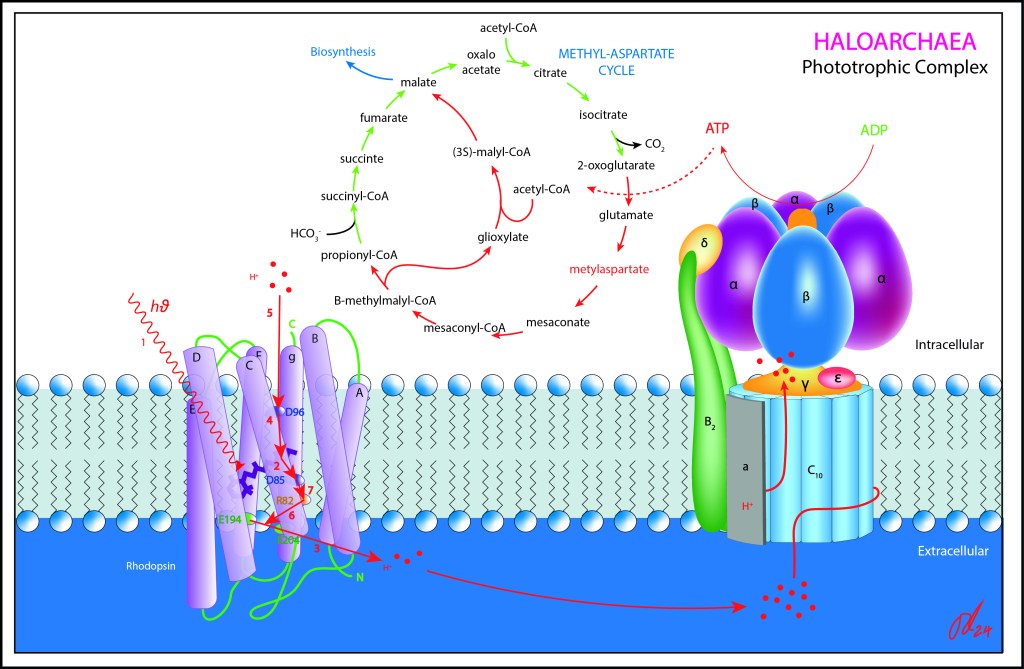
En arqueas, este complejo, denominado Bacteriorhodopsina BR, forma parte de un extenso arsenal de rhodopsinas presentes en arqueas, todas activadas por energía lumínica, grupo que incluye además las Halorhodopsinas HR (transporte hacia el citoplasma de iones Cl-) , Channelrhodopsinas CR (transporte Na+, K +, Ca2+, H+) , así como rhodopsinas sensoriales (SR) destinadas a orientar a las células hacia las fuentes de luz.
En general, todas las rhodopsinas son proteinas 7-transmembrana con retinal como cofactor, pero a diferencia del retinal presente en nuestros ojos, la luz isomeriza de un estado all-Trans a 13-Cis .
En BR, esta fotoisomerización genera a su vez un cambio conformacional al interior de la proteina, modificando la distribución de cargas eléctricas, particularmente en los aminoácidos D96, D85 , R82 y en el complejo Glu 204-Glu194 en la frontera externa de la proteína, convirtiendose estos elementos en receptores/dadores intermediarios del transporte de iones H+.
De esta manera, el nuevo estado eléctrico generado por lo isomerización del retinal, guía de manera controlada el transporte activo de protones desde el citoplasma hacia el exterior de la célula, generando finalmente un gradiente H+ (PMF), a ser aprovechado por la ATP syntasa.
El transporte de protones desde el citoplasma hacia el exterior de la célula no sigue una secuencia espacial dentro de la proteína, de manera que los movimientos parciales se dan “en desorden”, activandose primero D85 central (M1), luego PCR ubicado en frontera exterior, luego RSB y D96 interiores para finalmente concluir cerrando el ciclo R82 externo.
Este aparente desorden evita el tránsito iónico en sentido contrario, Back Propagation dirían los amantes de la inteligencia artificial, de los iones H+, optimizando así la eficiencia PMF.
Para garantizar la concentración iónica, el aparato fototrófico se ubica en invaginaciones de la membrana celular, lo que garantiza el confinamiento de carga H+ requerido para generar la diferencia de potencial a ser aprovechada por ATP Synthase .
Esta estrategia de confinamiento de cargas mediante invaginaciones de membrana se repite de manera casi que invariante en los diversos complejos fotosintéticos presentes en algas y plantas superiores, en membranas de bacterias fotosintéticas y también en los complejos fosforilativos presentes en las mitocondrias.
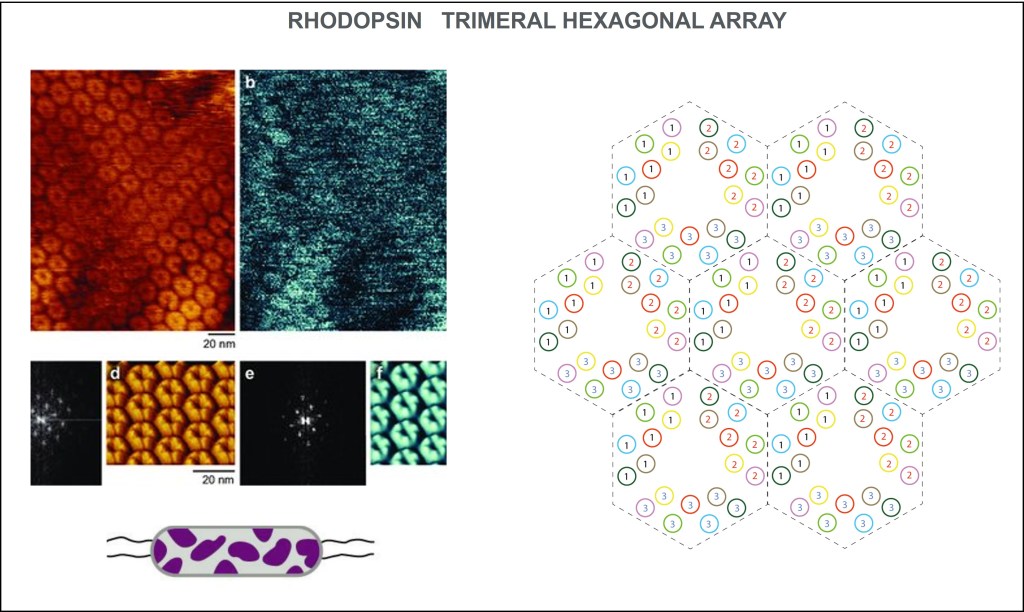
SIMETRÍA TRIGONAL-HEXAGONAL
La distribución de rhodopsinas en la membrana celular obedece a unos arreglos geométricos en tripletas moleculares conformando hexágonos a manera de panal, lo que optimiza el aprovechamiento de la superficie de membrana.
El proceso de aprovechamiento de luz para generación de PMF en arqueas es un proceso fototrofico, no fotosintético, lo que quiere decir que el carbono es incorporado al metabolismo celular a partir de moléculas parcialmente reducidas (HCO3-) y nunca (al menos en lo observado hasta el momento) a partir de CO2.
En arqueas, el ciclo del Metil-Aspartato es la ruta reductora preferente, utilizando como fuente de energía el ATP generado por la ATP Synthase, y generando malato como punto de partida para numerosas rutas de biosíntesis celular.
Orlando Rodríguez
Juno 2025
LECTURAS RECOMENDADAS
Evolución de las Arqueas
Adam PS, Borrel G, Brochier-Armanet C, Gribaldo S. The growing tree of Archaea: new perspectives on their diversity, evolution and ecology. ISME J. 2017 Nov;11(11):2407-2425. doi: 10.1038/ismej.2017.122. Epub 2017 Aug 4. PMID: 28777382; PMCID: PMC5649171.
Bacteriorhodopsinas
Yizhar O, Fenno L, Zhang F, Hegemann P, Diesseroth K. Microbial opsins: a family of single-component tools for optical control of neural activity. Cold Spring Harb Protoc. 2011 Mar 1;2011(3):top102. doi: 10.1101/pdb.top102. PMID: 21363959.
Ernst OP, Lodowski DT, Elstner M, Hegemann P, Brown LS, Kandori H. Microbial and animal rhodopsins: structures, functions, and molecular mechanisms. Chem Rev. 2014 Jan 8;114(1):126-63. doi: 10.1021/cr4003769. Epub 2013 Dec 23. PMID: 24364740; PMCID: PMC3979449.
Rhodopsinas sensoriales
Sasaki J, Takahashi H, Furutani Y, Kandori H, Spudich JL. Sensory rhodopsin-I as a bidirectional switch: opposite conformational changes from the same photoisomerization. Biophys J. 2011 May 4;100(9):2178-83. doi: 10.1016/j.bpj.2011.03.026. PMID: 21539785; PMCID: PMC3149258.
Para abordar el tema de la fotosíntesis, un buen punto de partida puede ser su historia evolutiva.
La fotosíntesis, entendida como la combinación del aprovechamiento directo de la energía lumínica y la incorporación de carbono en la maquinaria viviente a partir de CO2, solo se presenta en bacterias, algas y plantas superiores. En arqueas, la otra rama evolutiva de la vida, existe un mecanismo de captura directa de luz pero nunca incorporando carbono a partir de CO2.
En cuanto a las bacterias, la maquinaria fotosintética debió evolucionar a partir de transferencia horizontal desde maquinarias fosforilativas propias de bacterias heterótrofas.
No hay concenso en la comunicad científica en cuanto al origen evolutivo de la fotosíntesis; quizas los primeros pigmentos fotosintéticos fueron incorporados inicialmente como un mecanismo de protección UV (terrabacterias ancestrales colonizadoras de tierra firme con alta exposición al UV) , en un mecanismo similar a como son utilizadas las Xantofilas actuales, para luego ser aprovechados como elementos resonantes de la energía lumínica en vías similares a las actualmente utilizadas en la fotosíntesis.
Dos tipos de complejos fotosintéticos están presentes en los organismos fotosintétizadores: PSII, complejo áltamente oxidante, capaz de oxidar el agua, y PSI, muy buen reductor, capaz de reducir moléculas acumuladoras de energía.
Las maquinarias fotosintéticas en los diversos grupos evolutivos incluyen uno o los dos fotosistemas, PSII o PSI, encargados de la captura primaria de energía, y un complejo citocromo, BC1, análogo a B6f de los aparatos de fosforilación oxidativa, gestor de la fuerza protomotríz que luego será aprovechada por la ATP sintasa para generación de ATP.
Un salto evolutivo enorme lo dieron las cyanobacterias, quienes, seguramente por transferencia horizontal, se apropiaron y pusieron a operar de manera armónica los dos fotosistemas, con el citocromo compartido a manera de pivote del complejo fotosintético.
Complementaridad !!!!
Un elemento decisivo en la evolución de la vida.
Endosimbiosis a nivel bioquímico, lo que dió como resultado una sinergia de función entre PSI y PSII, siendo la evolución convergente mas impactante en la historia de la vida.
Surgen varios elementos de reflexión sobre el papel de la evolución divergente (Darwinismo) y la evolución convergente (Margulismo), y me atrevo a resumirlo en una frase:
“Darwin propone, Margulis dispone”.
Se requiere una oferta amplia de opciones evolutivas, proporcionada por las dogmáticas mutaciones al azar de Darwin, para que Margulis, independientemente de la indiscutible selección natural, tome opciones y las combine de manera complementaria en nuevos y más eficientes fenómenos simbióticos de alto impacto.
Darvin y Margulis permitieron entender la evolución ya no como ramas darwinistas que se desprenden de un único tronco común para competir y nunca más colaborar, tampoco raíces que de manera colabortiva convergen a dicho tronco, mucho menos raices tronco y ramas formando un “bonito” árbol de la vida, más bien una intrincada red de ramas y troncos que se separan, se unen en opciones cada más complejas, compiten, colaboran, luchan, sobreviven, fallan, se equivocan, desaparecen, comparten información, optimizan procesos, etc, formando una “red evolutiva” a manera de red neuronal gestora de la biodiversidad planetaria.
Así pues, los saltos evolutivos de mayor impacto se dan por mecanismos simbióticos, pero siempre requiriendo de una amplia oferta de opciones de evolución divergente.
Ensamblar una máquina altamente oxitadiva con una máquina fuertemente reductora es proporcionarle a la vida un motor turbocargado, capaz de oxidar el agua, un inagotable recurso energético, y así extraerle sus electrones, transportadores de la energía, y los protones, que luego aportarán a la fuerza protomotriz requerida por la ATP Sintasa , convirtiendo a sus portadores (cyanobacterias) en el gran protagonista de la trama de la vida hace 2,500 millones de años.
Y es que las cyanobacterias se convirtieron en EL JUGADOR (en mayúsculas), con el poder de “envenenar” la atmósfera con oxígeno, el inevitable desecho del agua, llevándose en su momento por delante el 95% de la vida en el planeta en un fenómeno conocido como Gran Evento de Oxidación (Huronian GOE), adquirieron el poder de actuar sobre la geología planetaria a gran escala, permitiendo que el hierro disuelto se oxidara de su forma ferrosa soluble a su forma férrica insoluble para generar las famosas bandas geológicas de hierro; poder para cambiarle el color a la tierra vista desde el espacio, de un rojizo propio de una atmósfera de metano y Co2 a un color azul propio del oxígeno, pasando en el medio por un color blanco al despojar al planeta de su abrigo de invierno (atmósfera de CO2 y Metano) convirtiendolo en una bola de nieve (Snow Ball), pero también aquiriendo el poder de ser punto de inflexión en la evolución de la vida.
Varios grupos de bacterias actuales operan con el fotosistema PSII, varios con PSI, pero fueron las cyanobacterias quienes aprovecharon el poder dual, generando una amenaza , …. y una oportunidad, para la vida en el planeta.
En respuesta a la amenaza, algunas arqueas Asgard y un linaje particular de proteobacterias aeróbicas, quizas aeróbicas facultativas, resolvieron la amenaza del oxígeno atmosférico en en un nuevo evento de simbiosis, primero con tímidos coqueteos sintróficos entre pares complementarios para llegar finalmente en una íntima unión que hoy conocemos como Eukariogénesis, células con núcleo.
A3 (Entagling, Engulfing Endosymbiosis) resume el fenómeno, que finalmente convierte al par simbiótico en organelos especializados , mitocontrias de orígen bacteriano, nucleos arqueanos.
De manera que cada célula de nuestro organismo viene a ser habitación de una arquea y una bacteria !!! . Somos en esencia bacterias y arqueas conviviendo en armonía !!!!
Y luego vino mas adelante un tercer evento simbiótico (otra vez Margulis, otra vez complementaridad), lo que permitiría incorporar una cyanobacteria (que luego se especializaria en forma de cloroplastos) al interior de la maquinaria eukariota, dando lugar a las algas, y mas adelante a las plantas superiores.