La “Fosforilación oxidativa” corresponde al proceso de captura de energía por parte de los organismos con respiración celular aerobia, incluyendo eucariotas (plantas, animales, hongos y protistas) y procariotas (bacterias y arqueas aerobias).
El término “Fosforilación oxidativa” resume de la mejor manera su función: “Fosforilación”, como objetivo, pues se trata de una maquinaria orientada a ACUMULARENERGÍA representada por radicales en ATP, y por otro lado “Oxidativa”, como requisito, pues la maquinaria opera mediante la oxidación (o extración de energía) de las coenzimas NADH y FDH2 procendentes del ciclo del Ácido Tricarboxílico (TCA) .
TCA a su vez opera como un intermediario en el tránsito de energía entre moléculas ricas en energía (carbohidratos y ácidos grasos) y la maquinaria fosforilativa.
PROCESO
La glucosa, donador principal de energía a la maquinaria fosforilativa, ingresa a la célula por dos vías:
1. A favor de gradiente (Difusión Facilitada) , sin requerirse ATP en el proceso, mediante transportadores de glucosa (GLUT).
2. En contra de gradiente (Trasporte Activo Secundario) mediante transportadores SGLT (Sodium-Glucose Linked Transporter), aprovechando las bombas Na⁺/K⁺ .
La familia de transportadores GLUT comprende 14 proteinas 12-transmembrana, GLUT1 a GLUT14, agrupadas en tres clases, todas con las terminales N y C expuestas al citoplasma.
El transportador GLUT4, en particular, es regulado por la insulina, cuya presencia en el torrente sanguineo inicia un proceso de transducción de señal que al completarse permite que GLUT-4 (almacenado en vesículas libres) se incorpore a la membrana celular para cumplir su función de transporte de glucosa hacia el citosol.
La regulación de glucosa por la insulina se sale de los objetivos de la presente lectura, pero definitivamente justifica un lectura aparte.
En el citosol, por la ruta de la glicólisis, cada molécula de glucosa es convertida en dos moléculas de Piruvato, las cuales ingresan a la mitocondria, también mediante difusión facilitada, con la participación de una proteína especializada presente en la membrana interna mitocondrial denominada MPC (Mitochondrial Pyruvate Carrier).
Finalmente, al interior de la mitocondria, el Piruvato cede su radical Acetil a la Coenzima A (CoA) mediante la enzima Piruvato-deshidrogenasa, lo que permite generar Acetil-Coenzima A (ACoA), molécula que activa el ciclo del ácido tricarboxílico o ciclo de Krebs. ACoA también es generado por radicales acil provenientes de la Beta-Oxidación de ácidos grasos, los cuales ingresan a la mitocondria en forma de Acil-Carnitina.
ACoA incorpora los dos carbonos del radical acil al oxaloacetato para regenerar citrato, lo que de paso define el nombre alterno del TCA (Ciclo del ácido cítrico).
TCA existe con el propósito fundamental de abastecer de combustible a la cadena fosforilativa en forma de FADH2 y NADH. Por cada molécula de glucosa se producen seis moléculas de NADH, dos de FADH2 y dos moléculas de GTP.
La maquinaria responsable de la fosforilación oxidativa (activada por NADH y FADH2) comprende cuatro complejos bioquímicos, numerados I a IV, los cuales tienen como función en conjunto generar un gradiente protónico entre la matriz y el espacio intermembrana de la mitocondria. Este gradiente a su vez alimenta la ATP Synthasa, lo que permite acumular energía en forma de ATP, de manera similar a como lo hacen los complejos fotofosforilativos en los organismos fotosintetizadores y a la fosforilación quimiosmótica propia de algunas bacterias y arqueas anaerobias (E. coli, desulfovibrio y arqueas metanógenas).
COMPELEJO I: NADH-UBIQUINONA OXIDOREDUCTASA
Encargado de la captura de energía almacenada en NADH, bombea cuatro iones H+ por cada molécula de NADH que ingresa al complejo. En el bombeo iónico participan numerosas moléculas de agua precisamente dispuestas al interior de las subunidades proteicas transmembrana PD y PP. (subunidades Distal y proximal transmembrana respectivamente), si bién su participación y manera de acción no está bién entendida.
El bombeo de protones contra gradiente es habilitado mediante una cadena de transporte de electrones (ETC) en el sentido NADH -> Ferredoxinas -> Ubiquinona , culminando con la reducción de la Ubiquinona y exportación de la misma en forma de Ubiquinol al pool de quinonas.
COMPLEJO II: SUCCINATO-UBIQUINONA OXIDOREDUCTASA
Es el único de los cuatro complejos que no realiza transporte de protones desde la matriz hacia el espacio intermembrana. De igual manera, único complejo intimamente Ligado al TCA, concretamente en la etapa de oxidación de Succinato a Fumarato, reduciendo FAD a FADH2, lo que activa una cadena de transporte de electrones en el sentido FADH2 -> Ferrredoxinas -> Heme -> Ubiquinona, las cuales finalmente son exportadas al pool de quinonas en su forma reducida.
COMPLEJO III: CITOCROMO bc1
Aceptor de las quinonas reducidas exportadas por los complejos I y II, captura los electrones del ubiquinol liberando 2 H+ por molécula los cuales se exportan al espacio intermembrana contribuyendo al gradente protónico (PMF).
Citocromo bc1 , al igual que b6f en los complejos fotosintéticos, enruta el primer electrón producto de la oxidación de la molécula de ubiquinol hacia una cadena de transporte de electrones compuesta por dos hemes BL -> BH para en el sitio Qi reducir parcialmente una molécula oxidada UQ2+ -> UQ+ . El segundo electrón es enrutado hacia el complejo Rieske proximal al espacio intermembrana, de allí a Cyt c1 y finalmente al cyt c soluble que se mueve hacia el complejo IV. El proceso reductor se repite para reducir completamente la ubiquinona parcialmente reducida UQ+ -> UQ2+ permitiendo regenerar en el pool de quinonas el primer ubiquinol extraído.
COMPLEJO IV: CITOCROMO C OXIDASA
Responsable del destino final de los electrones producto de las cadenas ETC de la maquinaria fosforilativa, produce agua a partir de O2 y H+ presentes en la matriz mitocondrial.
El complejo IV contribuye al PMF por dos vías: Por un lado consumiendo H+ en la matriz mitocondrial, y por otro lado transportando activamente H+ de la matriz hacia el espacio intermembrana, proceso mediado por su propia cadena ETC .
El complejo IV contiene dos centros de cobre:
Centro CuA: Contiene dos iones de cobre, siendo responsable de recibir los electrones provenientes de CytC, para luego transferirlos al centro catalítico CuB del complejo.
Centro CuB: Contiene un ión de cobre, que junto con un Heme a3 recibe los electrones de CuA para reducir completamente O2 a H2O evitando de paso que los electrones libres formen especies reactivas de oxígeno.
ATP SYNTHASE
Aunque técnicamente no forma parte del aparato fosforilativo, este complejo es el encargado de aprovechar el gradiente protónico para sintetizar ATP a partir de ADP, propósito único de la fosforilación oxidativa.
ATP Sintasa es una verdadera máquina ensambladora rotativa de tres tiempos: Admisión -> Compresión -> Expulsión, con sus centros activos separados 120° entre sí, utiliza para la rotación el gradiente protónico H+ generado por el aparato fosforilativo.
En algunas bacterias marinas anaerobias halófilas, propias de ambientes muy alcalinos, donde H+ no puede operar, ATP Sintasa utiliza de manera alternativa Na+ para generar la rotación requerida para la síntesis de ATP.
Antes de abordar el tema de pigmentos fotosintéticos, es conveniente revisar la estructura atómica y molecular del carbono.
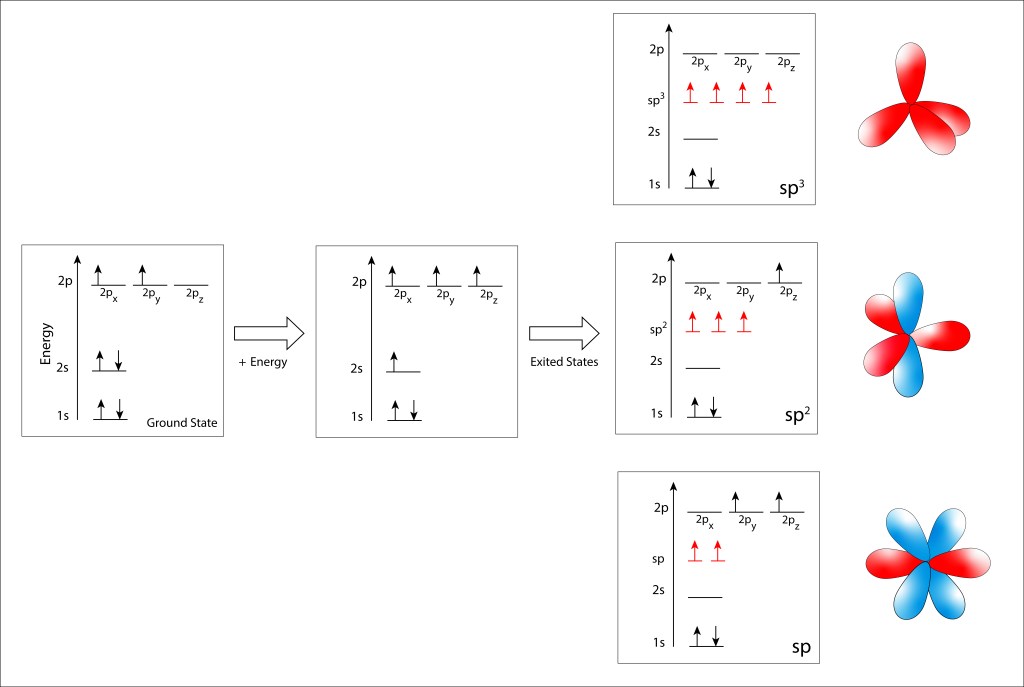
La Figura 1 ilustra de manera gráfica la distribución electrónica y los niveles de energía de los orbitales atómicos en el estado Ground y en los estados exitados sp3, sp2 y sp.
Estado exitado sp3 : Los cuatro electrones porvenientes de los orbitales 2s y 2p se hibridizan en un nivel equipotente de energía intermedio entre los niveles 2s y 2p, conduciendo a cuatro electrones sp3 separados espacialmente 109,5° formando un tetraedro regular.
Estado sp2 : Tres de los cuatro electrones se hibridizan de manera planar, separados 120° entre sí, quedando libre de hibridación el electrón 2pz cuya distribucion de probabilidad es perpendicular al plano sp3.
Estado sp: , Solo dos electrones se hibridizan y sus distribuciones de probabilidad se ubican sobre el eje x. En este caso, los dos electrones, 2py y 2pz se distribuyen a 90° entre sí en un plano perpendicular al eje x.
Figura 1. Orbitales atómicos Ground y exitados del átomo de carbono.
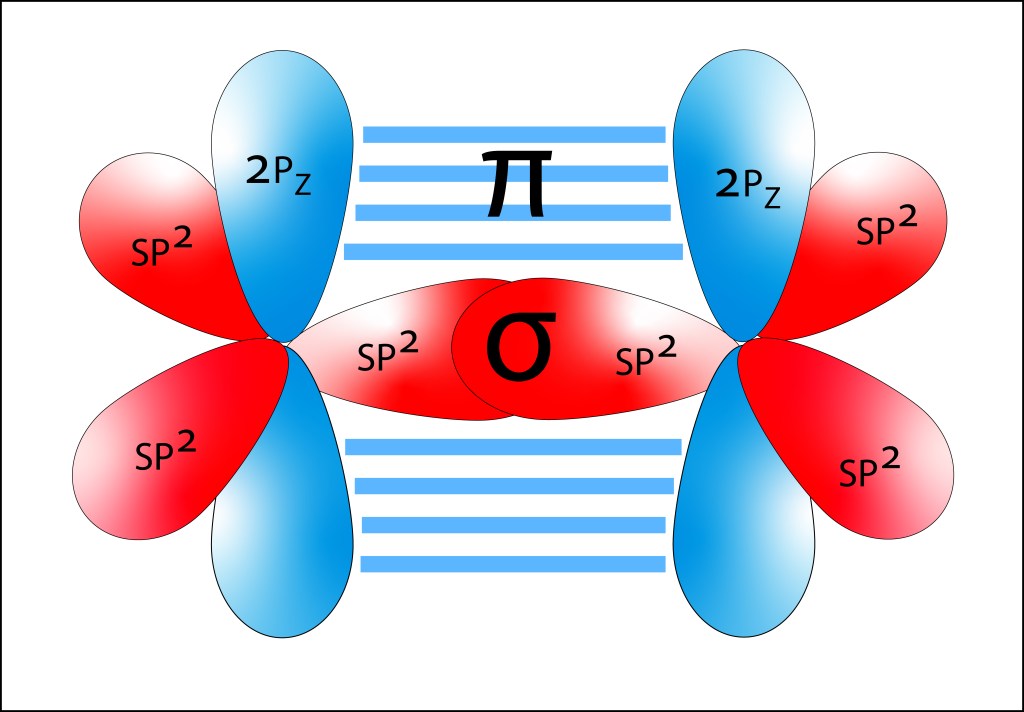
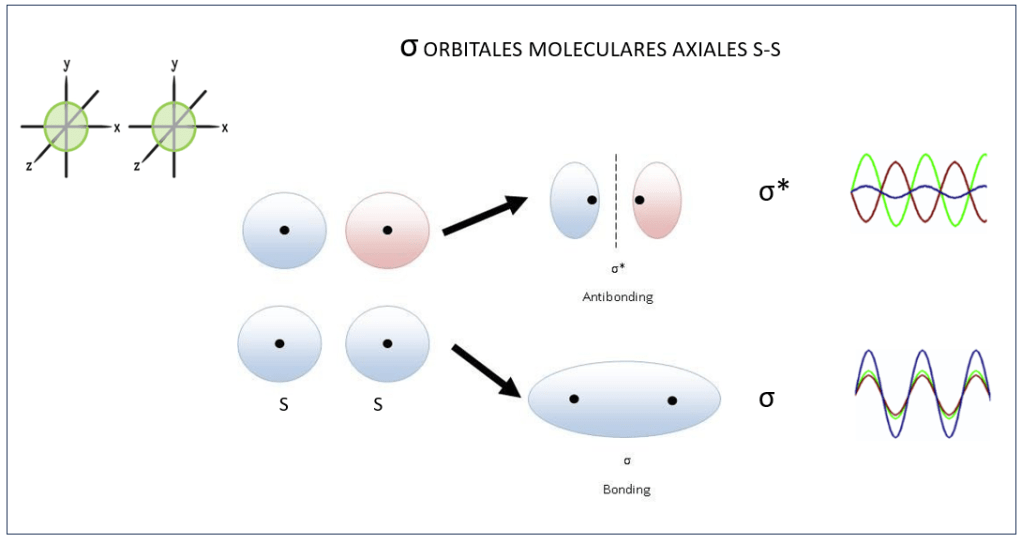
En los pigmentos fotosintéticos, Enlaces C=C alternan con enlaces C-C , sea de manera lineal (carotenos) o en anillo (clorofilas), formando lo que se denomina enlaces conjugados. Los enlaces C=C corresponden al estado exitado sp2 , en el que participan dos electrones: uno de los tres electrones híbridos sp2 y el electrón 2pz, ver Figura 2. Los dos electrones sp2 se alinean sobre un mismo eje, conformando un orbital molecular σ, mientras que los electrones 2pz se alinean de manera paralela, conformando un orbital molecular π.
Figura 2. Orbitales moleculares para un enlace C-C correspondiente a sp2
Dependiendo del estado de fase, dos tipos de enlaces σ pueden presentarse: Bond, con funciones de onda en fase, y Antibond, con las funciones de onda desfasadas 180° entre sí, ver Fig 3.
Figura 3. Orbitales moleculares σ y σ* , Bond y Antibond, dependiendo de los estados de fase de los electrones participantes..
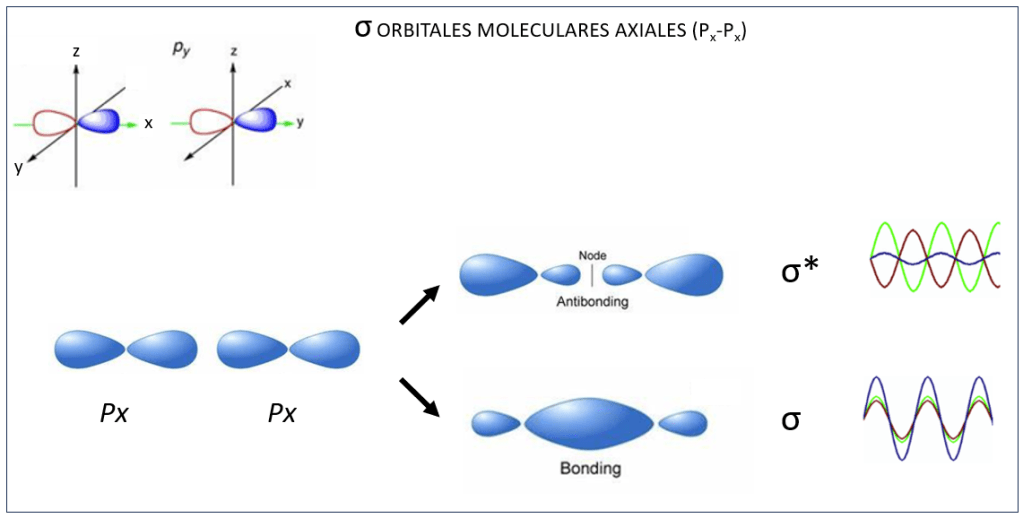
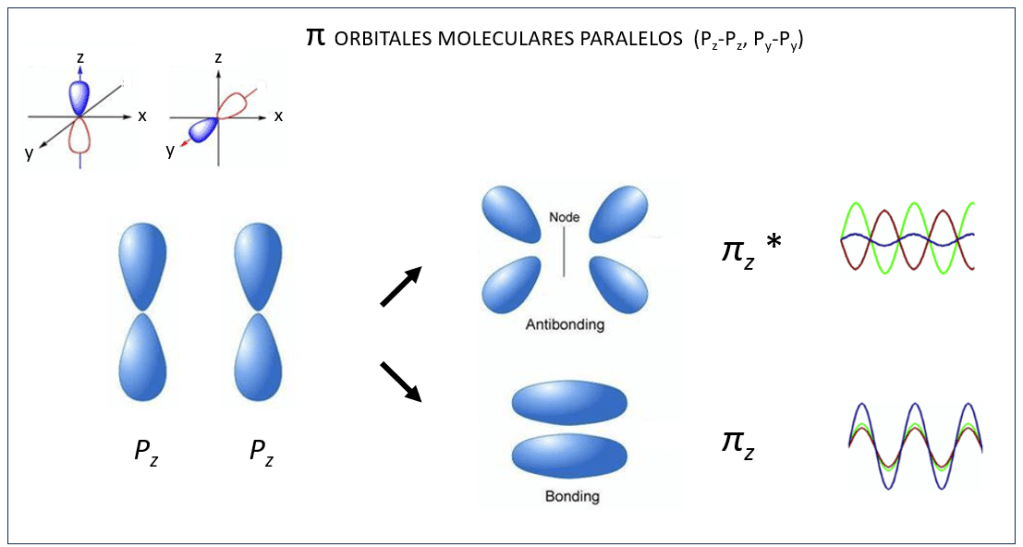
Orbitales moleculares tipo σ y σ* ocurren también entre orbitales atómicos px, ver Fig 4, mientras que py y pz interactuan de manera paralela conformando orbitales moleculares π y π*, ver Fig.5.
Figura 4. Orbitales moleculares σ y σ* entre orbitales atómicos Px
Figura 5. Orbitales moleculares π y π* entre orbitales atómicos py – py y pz-pz
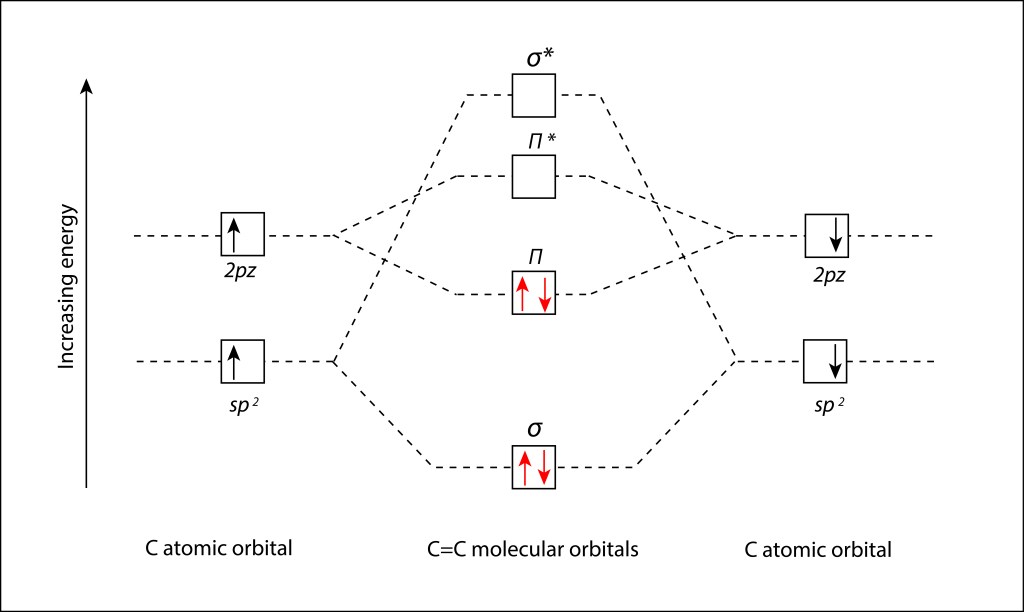
La figura 6 describe los niveles de energía de los orbitales moleculares correspondientes a los elementos del grupo 2 de la tabla periódica.
Figura 6. Niveles de energía de los Orbitales moleculares para los elementos del grupo 2 de la tabla periódica.
Partiendo de los niveles de orbitales moleculares especificados en la fig.6., se dpuede definir el diagrama orbital molecular (MO Diagram) correspondiente al enlace C=C presente en los enlaces conjugados de los pigmentos fotosintéticos; ver Fig 7.
Figura 7. Diagrama orbital molecular (MO Diagram) para el enlace C=C en estado Ground
HOMO – LUMO
Los términos HOMO / LUMO hacen referencia a los orbitales “Highest Occupied Molecular Orbital” y “Lowest Unoccupied Molecular Orbital”, que, para el caso del enlace C=C especificado en la Fig.7., corresponden a los orbitales moleculares π y π*, respectivamente.
Se mencionó anteriormente que el orbital molecular π es un orbital “Bonding” en el cual las funciones de onda de los electrones que participan están en fase, ver Fig.5., mientras que π* es un enlace “Antibonding” , en desfase de 180°.
Cuando un fotón incide sobre un electrón π con un energía exactamente igual a la diferencia entre los niveles de energía (π* – π), en electrón se exita pasando del nivel π al π* , esto es, de HOMO a LUMO, ver Figura 8.
Figura 8. Diagrama orbital molecular (MO Diagram) para el enlace C=C en estado exitado
Los diagramas propuestos por Richard Feynman describen de una manera elegante la interacción fotón-electron, aplicable a los pigmentos fotosintéticos, ver Fig 9.
Figura 9. Diagrama de Feynman correspondiente a l salto de energía de un electrón π ground (HOMO) a su estado exitado π* (LUMO)
La exitación electrónica HOMO-LUMO de π a π* es el fenómeno natural al cual la vida a travez de la fotosíntesis ha recurrido para aprovechar la energía solar, con el único propósito de mantener su estructura y función orgánica, lo que viene a ser el motor mismo de la vida en nuestro planeta.
ESTADOS TRIPLETE Y SINGLETE
En adición a la energía de exitación inherente al tránsito HOMO-LUMO del electrón (π -> π*) , el momento angular del electron, denotado por S, o Spín, juega igualmente un rol importante en la captura de energía y relajación en los procesos fotosintéticos.
El Spin es un número cuántico inherente a las partículas que conforman el universo. En el caso de los fermiones, a los cuales pertenece el electrón, su valor siempre es S=1/2, y su proyección sobre el eje z, o sz , puede tomar los valores +1/2 (up ↑) y -1/2 (down ↓).
Matemáticamente,
Los dos eigenvalues de Sz, , corresponden a los siguientes eigenspinors:
Para un sistema conformado por dos electrones, hay dos estados de importancia en los procesos fotosintéticos:
Estado triplete: Comprende tres estados antisimetricos con spin total S=1
Componentes: ms=+1 , ms = 0 , ms −1, donde
∣T1⟩=∣↑↑⟩
∣T0⟩=1/√2 (∣↑↓⟩+∣↓↑⟩)
∣T−1⟩=∣↓↓⟩
Estado singlete (S = 0): Estado simétrico con spin total S=0 .
Componente único : ms=0, donde
∣S⟩=1/√2(∣↑↓⟩−∣↓↑⟩)
Por regla general, para dos electrones en el mismo sistema, el estado singlete (estado simétrico) suele tener mayor energía que el estado triplete, debido principalemente a que un estado simétrico permite mayor cercanía entre el par de electrones que en estados antisimétricos, lo que aumenta la repulsión coloumbiana.
En la fotosíntesis y otros fenómenos bioquímicos, una e las estrategias de de-exitación se conoce como Inter-System Crossing, ISC. Electrones pueden pasar de estados Singlete de alta energía a estados triplete de menor energía, ver Fig. 10, como un paso intermedio para retorno a GND.
Figura 10. IC – ISC como rutas de De-exitación no radativa.
ENLACES CONJUGADOS
Como se mencionó, cadenas y anillos con alternancia de enlaces simples ( σ) y dobles (σπ) se denominan enlaces conjugados. ver Fig.11. En este tipo de arreglos, los electrones π se deslocalizan, lo que otorga una gran estabilidad a la molécula a mas de ser fenómeno base de sus propiedades resonantes.
La diferencia de energía (π* – π) en los pigmentos fotosintéticos depende de numerosos factores, intramoleculares e intermoleculares. Los enlaces conjugados, como parte de la estructura molecular, definen en buena medida el valor de dicha diferencia .
Figura 11. Estructura molecular de la porfirina, con un anillo conformado por 9 enlaces σπ alternando con 9 enlaces σ.
A mayor número de enlaces conjugados por pigmento menor la distancia energética (π* – π), lo que se traduce, en pigmentos fotosintéticos, a absorber fotones a menores energías, tan bajas como el infrarrojo cercano.
Cada grupo biológico dependiente de la fotosíntesis tiene su entorno característico, con una luminosidad característica, lo que implica que la maquinaria fotosintética debe sintonizarse con la fuente de energía disponible. El ajuste de la diferencia HOMO-LUMO es la herramienta base para dicha sintonización.
No quisiera cerrar esta lectura sin mencionar a un personaje que de una u otra manera entendió la importancia de la luz en el universo.
Me refiero al faraón Akhenaton, uno de los últimos faraones de la dinastía XVIII, quien en medio de un gobierno teocrático politeista se atrevió a implementar, por primera vez en la historia de la humanidad, el monoteismo en su reino. Así, el culto a Ra, el dios sol, se convirtió en el centro de la religión durante su regencia . Esta “herejía”, opuesta al zoológico de dioses defendidos por los sacerdotes de la época, marcó el principio del fín de su dinastía, quedando sin embargo “grabado en piedra” para la posteridad su aporte al reconocimiento del papel de la energía electromagnética, entiendase Ra, sol, luz o fotón, en la vida y evolución de nuestro planeta.
Obviamente, tras Akhenaton vinieron aportes menos religiosos y más científicos, dentro de los que cabe resaltar los aportes de J.C. Maxwell, Max Plank , Albert Einstein y Richard Feynman, entre otros, pero eso debería ser tema para otras lecturas.
Figura 1. Arreglo de pigmentos de una subunidad tubular en el cuerpo del Clorosoma de Bacterias verde sulfurosas
El clorosoma es el organelo responsable de la captura de energía lumínica en las maquinarias fotosintéticas de tres grupos de bacterias Gram (-) :
Bacterias Verdes Sulfurosas (GSB), representadas por el género Chlorobium ( C. tepidum), y los géneros Chlorobaculum, Prosthecocloris y Cloroherpeton.
Bacterias Fotótrofas Anoxigénicas Filamentosas (FAP), anteriormente bacterias verdes no sulfurosas, representadas por el género Chloroflexus (C. aurantiacus), y los géneros Roseiflexus, Oscilochloris, Heliothrix y Chloronema.
Acidobacterias, con un único género fotosintético descrito a la fecha, Chloroacidobacterium.
Equivalente en función a los complejos LHII y LHI de bacterias púrpura y a los Ficobilisomas de las Cyanobacterias, los clorosomas son los orgenelos responsables de capturar la energía lumínica y redireccionarla al centro de reacción RC donde la separación de cargas tiene lugar.
Los Clorosomas se diferencian de los demás complejos antena por su gran tamaño (formacilindrica de corte elipsoidal con una longitud entre 100 y 200 nm, 50 nm de anchura, 25 nm de altura), por la ausencia de una matriz proteica que de soporte y organización a los pigmentos fotosintéticos y por el elevado número de pigmentos por unidad (250,000 moléculas de bacterioclorofilas c, d y e por clorosoma).
En ausencia de la matriz proteica, los pigmentos se autoensamblan en agregados paracristalinos, ver Fig.1., lo que los hace objeto actual de investigación en nanotecnología (fotosíntesis artificial y electrónica molecular).
El compacto agregado de pigmentos fotosintéticos proporciona un acoplamiento exitónico intramolecular muy fuerte. Este elemento, sumado a la enorme cantidad de moléculas involucradas, son la base de la alta eficiencia en captura de energía lumínica, al punto que unos pocos fotones incidentes por pigmento por día le permiten a las bacterias prosperar en ambientes anóxicos de extremada baja luminosidad (hasta 100 metros de profundidad en el Mar Negro), siendo considerados a la fecha los más eficientes complejos antena encontrados en la naturaleza.
ANATOMÍA DEL CLOROSOMA
El clorosoma está conformado por un cuerpo, responsable directo de la captura de energía, una capa conectora base (Baseplate), y en el caso de GSB, una capa proteica especial, FMO, (Fenna-Matthews-Olson), responsable de transmitir la energía al centro de reacción RC, ver Fig.2.
Figura 2. Estructura general de un clorosoma
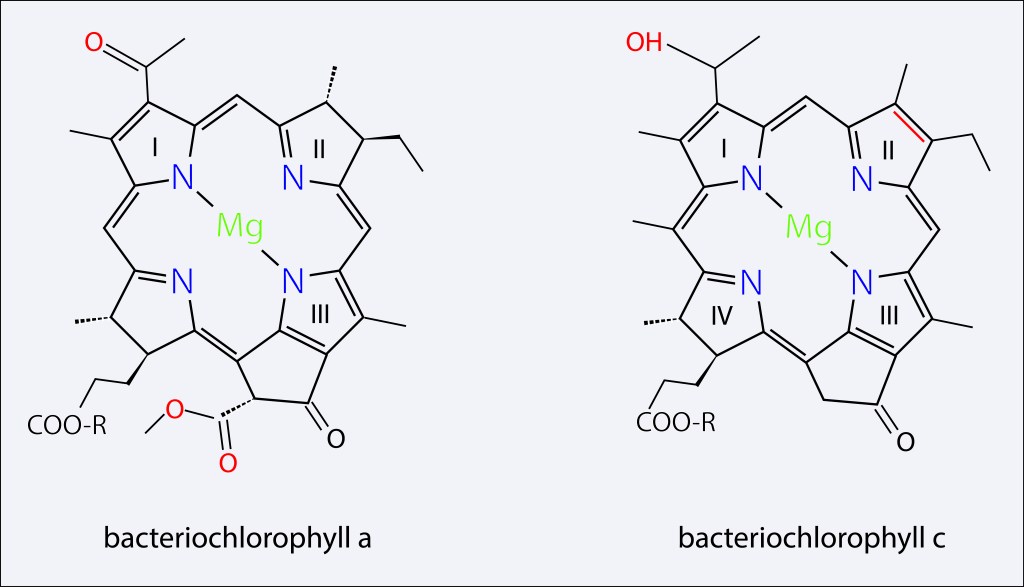
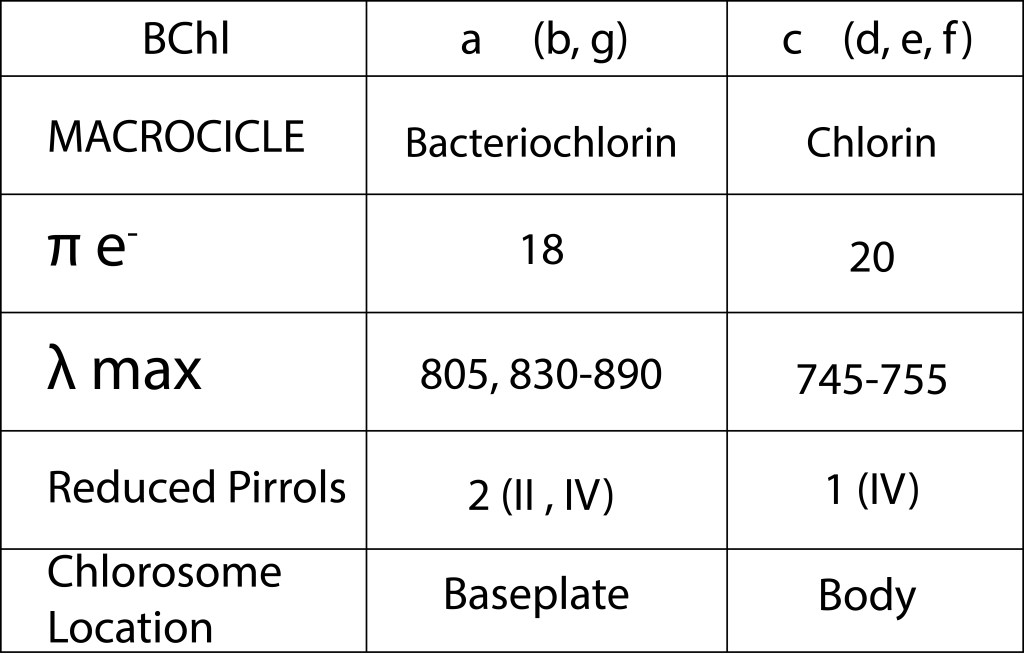
Los dos principales pigmentos presentes en el clorosoma corresponden a BChl c y BChl a , en cuerpo y baseplate respectivamente, ambos con un precursor común, Chlorophyllide a, pero con diferencias importantes en su estructura molecular como se ilustra en la fig 3 y en la tabla 1.
Figura 3. Pigmentos presentes en el clorosoma
Tabla 1. Características de los pigmentos mas representativos presentes en el clorosoma
Los pigmentos fotosintéticos responsables de la captura primaria de energía corresponden principalmente a bacterioclorofilas del tipo CBhl c, en un número entre 60,000 y 250,000 unidades por clorosoma, cantidad que depende de la especie y de la disponibilidad de energía lumínica del entorno. Chlorobium tepidum contiene entre 200 y 250 clorosomas, para un total de cerca de 50 millones de moléculas de BChl c, lo que equivale a cerca del 30% del carbono celular.
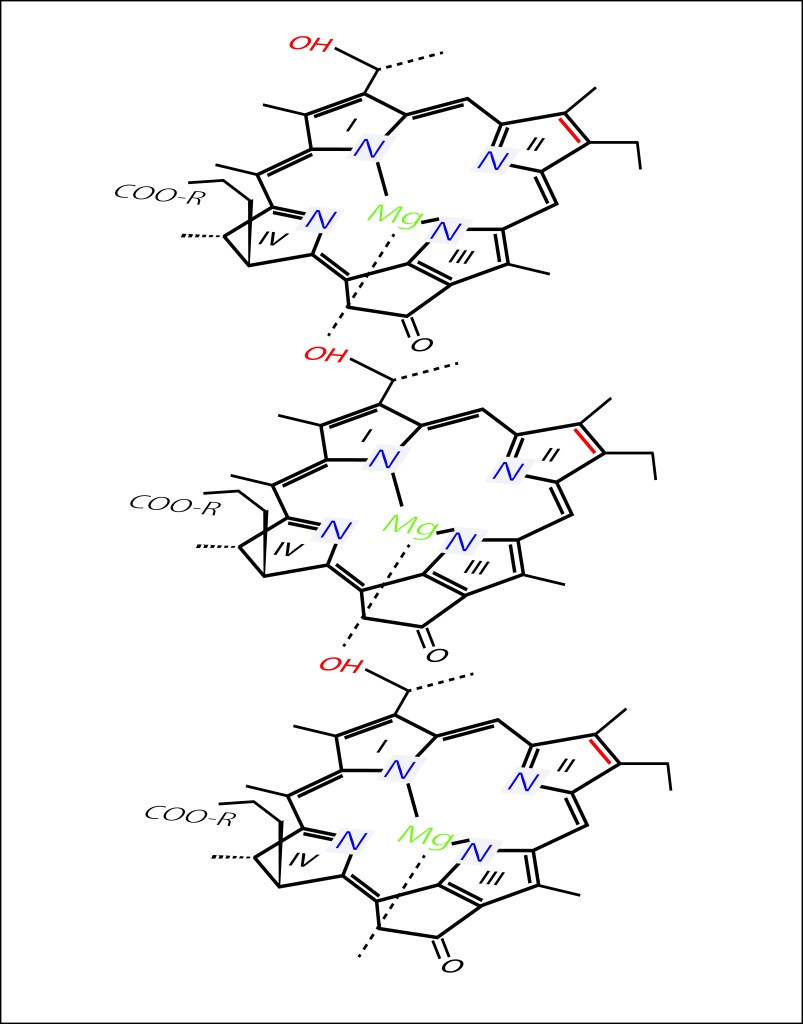
Al interior del cuerpo los pigmentos se autoensamblan en subunidades tubulares, ver Fig1., mediante enlaces de coordinación C-31OH-Mg, ver Fig 4; absorben energía a 750 nm y por resonancia de Förster y coherencia cuántica direccionan la misma a la Base plate a 795 nm .
Figura 3. Apilamiento de moléculas BChl C bajo una estructura “Parallel Stack”. Otros arreglos son posibles (Piggy-back dimer model, Anti-parallel monomer stack model y Syn-anti monomer stack model). Visitar: Photosynthesis Research · June 2013. DOI: 10.1007/s11120-013-9869-3 · Source: PubMed. Para todos los casos, un enlace de coordinación C-31OH-Mg es el elemento enlazante de la cadena. Para los demás arreglos,
El conjunto de tubulos está rodeado por una membrana monofosfolipídica de geometría cilíndrica y corte elipsoidal poblada de proteinas clorosómicas transmembrana (CSM) pertenecientes a las familias CsmA, CsmB/CsmF, CsmC/CsmD y CsmH/I,J,X. La función de estas proteínas es contribuir a la estabilización y organización estructural de los pigmentos, y en el caso de CSMJ protejer el sistema mediante disipación de energía lumínica (quenching ).
En adición a las bacterioclorofilas, el cuerpo contiene carotenos que participan en la captura y transmisión de energía, y quinonas , particularmente clorobiumquinona, que operan como agentes protectores quencher.
En cuanto a la capa basal del clorosoma, o Base Plate, está conformada por proteínas CSMA ligadas a la capa monofosfolipídica en una estructura paracristalina 2D. las proteínas a su vez contienen pigmentos BChl a y probablemente carotenos. Cada monómero de CSMA está compuesto por dos hélices alpha, donde la hélice N-Terminal (residuos 6-36) se encuentra inmersa en los lípidos mientras la hélice C-Terminal (residuos 41-49) se asocia a la proteina FMO.
Hélices Alpha de monómeros adjuntos establecen interacciones hidrofóbicas que favorecen conformaciones diméricas, las cuales son la base del arreglo molecular de la Base Plate.
Finalmente, la capa FMO, que conecta la Baseplate con en centro de reacción en bacterias verde sulfurosas, es un trímero proteico con simetría de orden 3, compuesto por monómeros idénticos, ver Fig 4.
Cada monomero de CSMA contiene en su interior siete bacterioclorofilas BChl a, ver Fig 5, aunque en Chlorobaculum tepidum y otras bacterias verdes sulfurosas se ha encontrado evidencia de una octava BChl a más débilmente ligada, que parece cumplir un papel de acoplamiento con la Baseplate. Estos pigmentos son los responsables finales del tránsito de energía desde el complejo antena hasta el centro de reacción.
Figura 5. Disposición de las BChl a al interior de los monómeros CSMA. Fuente: Energy-scales convergence for optimal and robust quantum transport in photosynthetic complexes. The Journal of Chemical Physics · April 2011. DOI: 10.1063/1.4856795
Para concluir, es de anotar que FMO fué el primer complejo proteina-pigmentos en ser estudiado y entendido. Gran parte de lo que se conoce hoy en día de la interacción entre pigmentos y proteinas proviene de los estudios de dicho complejo. FMO fué igualmente el primer caso en que se demostró la ocurrencia de coherencia cuántica como fenómeno fundamental en la transmisión de energía hacia el centro de reacción. FMO solo está presente en especies con centros de reacción tipo I, carcterizados por la presencia de ferredoxina como aceptor de electrones en la cadena de transporte de electrones ETC, tema que se tratará en las próximas lecturas.
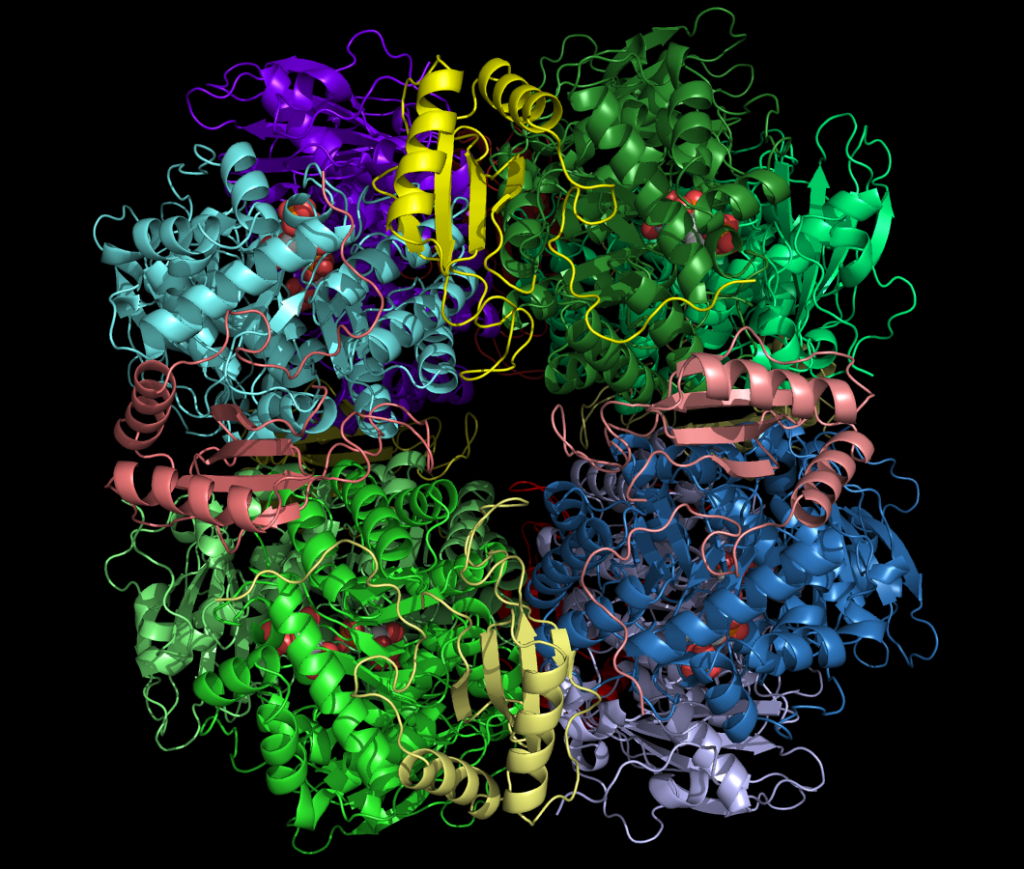
Figura 1. Vista 3D de la enzima Rubisco, responsable de la fijación de carbono en el ciclo de Calvin-Benson.
El ciclo de Calvin-Benson es de lejos el ciclo reductor mas importante en la naturaleza, presente en varios grupos bacterianos, algas y plantas superiores.
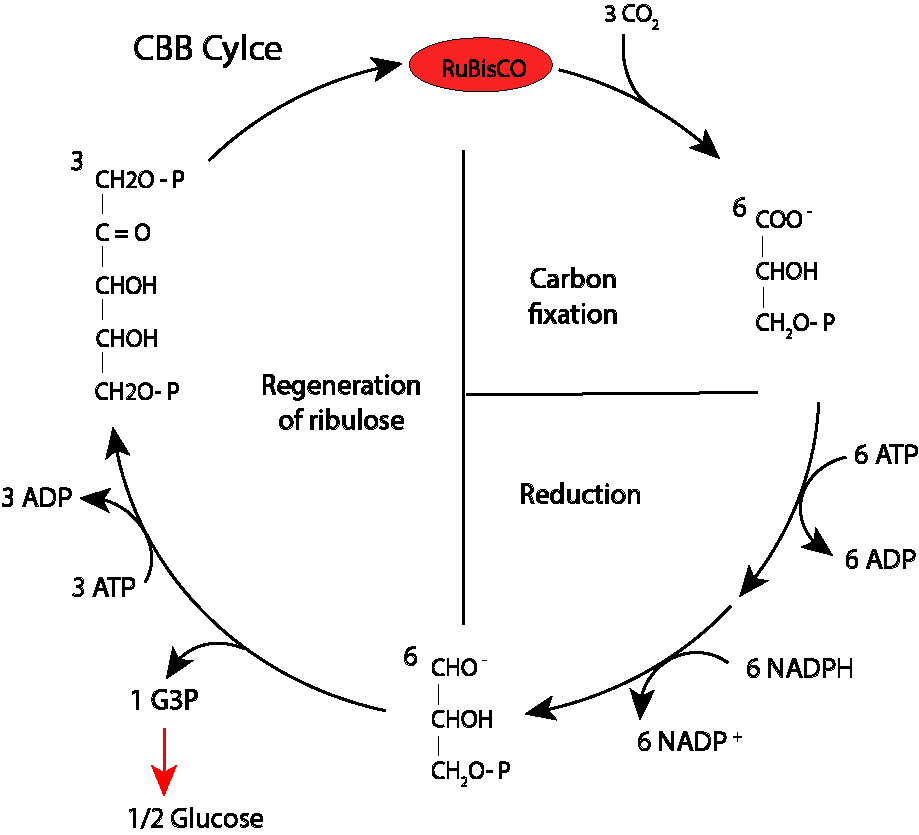
La manera más común de representar el ciclo se ilustra en la Fig 2.
Figura 2. Ilustración general del ciclo de Calvin-Benson
Varios elementos a resaltar:
Se trata de un ciclo reductor, responsable de la biosíntesis a partir de Gliceraldehido 3 fosfato (G3P) , mediante incorporación de CO2, ruta característica y propia de los procesos fotosintéticos.
El ciclo requiere energía externa, en forma de ATP y NADPH
El ciclo comprende tres fases: Carboxilación, Reducción y Regeneración
Durante la fase de carboxilación, 3 moléculas de CO2 son incorporadas al ciclo, y en conjunto con 3 moléculas de H2O y tres de Ribulosa 1-5 bifosfato se producen seis moléculas de 3-fosfoglicerato. En esta fase la enzima Rubisco (Ribulosa 1-5 bifosfato carboxilasa oxigenasa) es la responsable de la carboxilación.
En la fase reductora, seis moléculas de Gliceraldehido 3-fosfato (G3P) son sintetizadas a partir de igual número de moléculas de 1-3 bifosfoglicerato.
Finalmente, en la fase de regeneración, cinco de las seis G3P producidas previamente se destinan a la regeneración de 3 moléculas de Ribulosa 1-5 bifosfato, la sexta G3P es extrída del clico para biosintesis o como reserva de energía, propósito fundamental del proceso.
Lo que la Fig 2. no muestra es la complejidad de la fase de regeneración.
Para ilustrar de la mejor manera esta fase resulta mucho mejor “abrir” el ciclo, ver fig 3., con tres moléculas de Ru-1-5-BP iniciando la ruta metabólica y “OTRAS” tres finalizandolo, lo cual de paso es la manera mas estricta de representar el proceso. El hecho de que la ruta se repita ciclicamente no quiere decir que sea circular, lo que implicaría las que las mismas moléculas de Ru-1-5 BP son las que incician y cierran el ciclo.
Como se mencionó arriba, una de las seis moléculas de G3P producidas durante la fase de reducción se destina a biosintesis (o como reserva de energía), mientras que las cinco restantes se destinan a la regeneración de Ru-1-5BP.
Figura 3. Representación alterna de la Ruta metabólica de Calvin-Benson
Se incluyen en la parte superior de la figura las dos rutas de biosíntesis más relevantes: La biosíntesis de almidón (en el cloroplasto de plantas superiores) y la biosíntesis de sucrosa (en el cytosol).
Para facilitar la comprensión del proceso, moléculas orgánicas de tres carbonos se representan como triángulos, de cuatro como cuadrados, de cinco como pentágonos, etc.
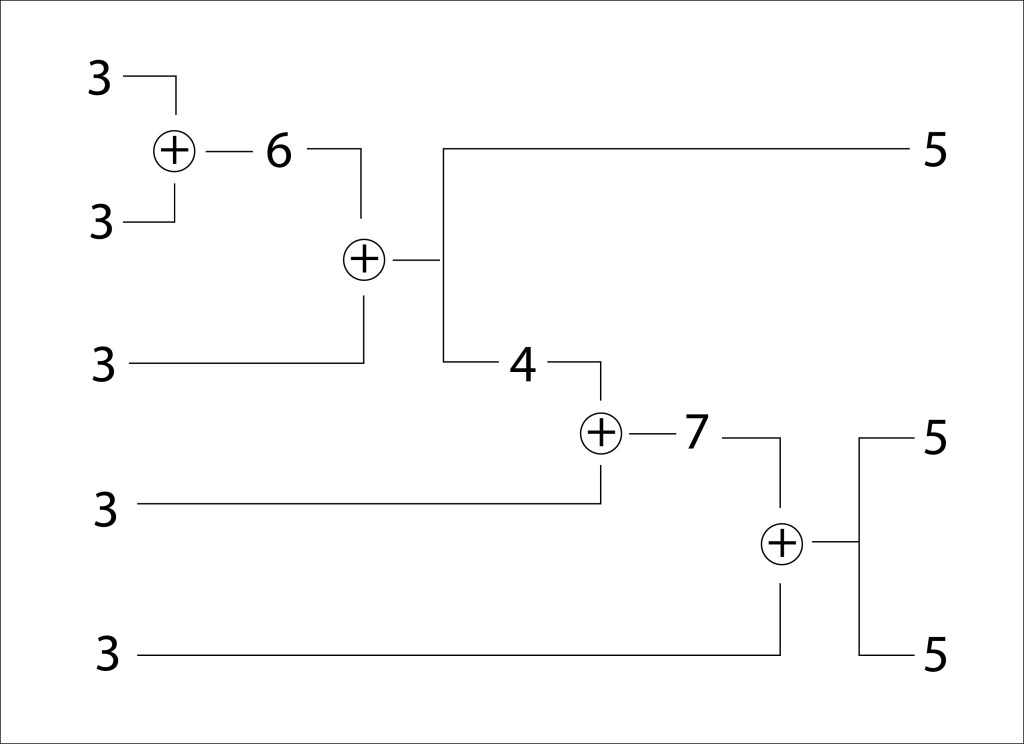
La aritmética implicada en el reagrupamiento de carbonos en la fase de regeneración, esto es, la síntesis de tres moléculas de cinco carbonos (Ribulosa 2-5 bifosfato) a partir de cinco moléculas de tres carbonos (Gliceraldehido tres fosfato) se resume en la fig 4.
Figura 4. Aritmética de la fase de regeneración en el ciclo de Calvin-Benson
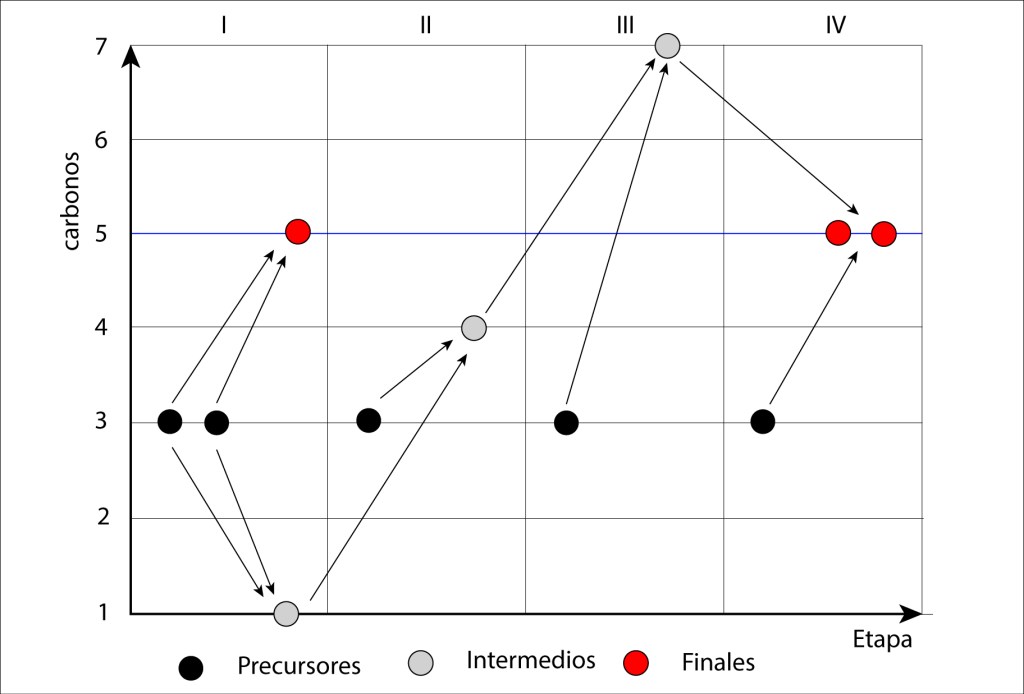
Otra manera de visualizar la fase de regeneración es mediante un diagrama de evolución temporal en el número de carbonos/molécula, ver Fig 5.
Figura 5. Evolución de carbonos a lo largo de la fase de regeneración
Lo que permite analizar este gráfico es la distribución de densidad de carbonos por molécula alrededor del objetivo final (5 carbonos).
Una manera de cuantificar esta densidad es a travez de un índice de variacion I, de acuerdo a la fórmula especificada en la figura. Para este caso, 2,36 es el valor calculado del índice de densidad de carbonos, teniendo en cuenta las moléculas precursoras de tres carbonos, G3P, moléculas intermedias de 4, 6 y 7 carbonos, ver fig.3, y las moléculas objetivo, Ru-1-5 BP.
No existe una fase de regeneración alterna que aporte un úndice inferior a 2,36. El número de carbonos oscila entre 3 y 7 carbonos, cualquier alternativa con moléculas de menos de tres carbonos o más de siete termina arrojando un índice mayor.
A manera de ejemplo, analizemos una ruta alterna como la especificada en la fig. 6.
Figura 6. Aritmética alterna de carbonos a lo largo de la fase de regeneración
En este caso, en lugar de la síntesis de F1-6BP (fructuosa bifosfato) a partir de G3P y DHP, con alguna enzima podría generarse X5P precursor de Ru1-5BP y una molécula orgánica de un carbono, que fosforilada convenientemente (inversión en energía), podría integrarse a un G3P para formar E4P .
Lo demás quedaría igual.
Esta modificación metabólica en la fase de regeneración termina generando cambios significativos en la densidad de carbonos, ver fig.7., como lo demuestra el nuevo índice de densidad calculado I=3,73 vs I=2,36 en el caso real.
Figura 7. Evolución alterna de carbonos a lo largo de la fase de regeneración
Con este ejercicio la pregunta que surge es, cuál es el significado del índice de densidad de carbonos?
En primer lugar, la energía invertida en cualquier otra ruta de regeneración alterna debe incrementarse,en el caso presentado por la integración de moléculas de un carbono a G3P, cuya activación por fosforilación implica inversión adicional de ATP, pero también por la energía invertida en la síntesis de enzimas adicionales para habilitar la ruta alterna.
Teniendo en consideración que la vida y su fenomenología bioquímica no son ni pueden ser una excepción a las reglas del universo, lo que se está midiendo es el costo metabólico de las rutas metabólicas, correspondiendo el menor costo, menor esfuerzo o mínima acción, a la ruta seleccionada por la naturaleza, lo que debe tener un equivalente discreto de la minima acción de la mecánica lagrangiana aplicada a fenómenos coninuos, acción que finalmente es la medida de la evolución temporal de las energías cinéticas y potenciales del sistema.
Las bacterias púrpura son un grupo de proteobacterias Gram negativas pertenecientes al órden Chromatiales, clase Gammaproteobacterias, presentes en cuerpos de agua anóxicos con alta iluminación.
A nivel bioquímico es un grupo muy versátil, pues dependiendo de las condiciones y recursos energéticos disponibles puede activar rutas de metabolismo fotoautótrofo, fotoheterótrofo, fermentativo, o respiratorio (anaerobio o aerobio).
Dada la importancia ecológica del grupo (particularmente como agente purificador de aguas residuales), su bioquímica ha sido ampliamente estudiada. La comprensión de su función fotosintética es de gran ayuda para abordar la dinámica de maquinarias más complejas, particularmente las correspondientes a cyanobacterias, algas y plantas superiores.
Antes de describir en detalle dicha maquinaria, vale la pena revisar algunos conceptos generales del fenómeno.
La fotosíntesis se define como el proceso de captura de energía lumínica y transpormación a energíia química, utilizando CO2 como fuente de carbono, estando presente en varios grupos bacterianos de bacterias, en algas y en plantas superiores.
El proceso comprende dos fases:
REACCIONES DEPENDIENTES DE LA LUZ: Ejecutadas de manera secuencial por diferentes complejos proteínicos transmembrana, parten de la captura de energía lumínica y se extienden hasta la síntesis de ATP y NADPH, ver Fig.1.
Fig.1. Etapas de las reacciones dependientes de la luz en la fotosíntesis.
REACCIONES NO DEPENDIENTES DE LUZ: Corresponde a un conjunto de reacciones de biosíntesis partiendo de agua y CO2, utilizando ATP y NADPH como fuentes e energía. Esta fase se realiza en el citoplasma mediante diversas rutas metabólicas reductoras, que dependiendo del grupo biológico pueden corresponder al ciclo de Calvin-Benson-Bassham (ciclo de las pentosas fosfato reductoras) , al ciclo reverso del ácido tricarboxílico o a la ruta del hidroxi-propionato, principalmente.
REACCIONES DEPENDIENTES DE LA LUZ
En las bacterias púrpura, las reacciones fotosintéticas dependientes de la luz ocurren en organelos especializados de la membrana interna celular, denominadas cromóforos o cromatóforos, ver Fig 2.
Fig 2. Cromatóforo en Rhodobacter sphaeroides. Fuente: Sener MK et al, 2007. Atomic level structure and function of an entire biological membrane: the photosynthetic unit of Rhodobacter sphaeroides. Proc Natl Acad Sci USA 104
Los cromatóforos surgen de invaginaciones de la membrana citoplasmática interna. Cada invaginación genera un volumen periplasmático confinado, limitado hacia el exterior por la membrana externa, ver Fig.3.
El reducido volumen de periplasma al interior del cromatóforo optimiza la generación de un gradiente de carga positiva, PMF, esencial para la síntesis de ATP.
Estrategias de confinamiento equivalentes se presentan en los Tilakoides de los cloroplastos, las invaginaciones tilakoidales de cyanobacterias, en el espacio periplasmático de heliobacterias e incluso en las crestas mitocondriales propias de células eukariotas.
Fig 3. Estructura de un cromatóforo
Las reacciones dependientes de la luz comprenden las siguientes etapas:
CAPTURA DE LUZ
EXITACIÓN DEL PAR ESPECIAL DE CLOROFILAS PRESENTES EN EL CENTRO DE REACCIÓN RC TIPO II (P870)
FLUJO DE ELECTRONES (Electron Transport Chain, ETC) a lo largo de una sucesión de reacciones de oxidoreducción a partir de la exitación y oxidación del par especial de clorofilas.
FUERZA PROTOMOTRIZ PMF (generación de un potencial protónico)
SÍNTESIS DE ATP (a partir de PMF como energía generatriz).
La Fig.4. resume a manera de diagrama de flujo el proceso fotosintético en este grupo, resaltando el colores los diferentes tiposde energía, incluyendo en rojo la energía lumínica (correspondiente a la captura y transporte de fotones), en verde la energía eléctrica, cuyo flujo se denomina ETC (Electron Transport Chain), en amarillo el flujo protónico (o flujo de iones H+) y finalmente en negro la depositación final de energía en la forma de ATP. Se omite en el diagrama la síntesis de NADPH, tema que se abordará más adelante, el cual requiere ATP y enzimas reductoras específicas. Con fines ilustrativos el centro de reacción RC y el complejo II se tratan como bloques independientes, si bién corresponden a un solo heterodimérico con sumbunidades L, M.
Fig 4. Diagrama de flujo reacciones fotosintéticas depenientes de la luz en bacterias púrpura
Como se aprecia en la figura, el complejo fotosintético de este grupo bacteriano se caracteriza por tener un solo fotosistema (Tipo II) , el cual es responsable de la reducción de quinonas y depositación de las mismas en el pool de quinonas, y un citocromo BC1 que activado por las quinonas reducidas responde por la generación de fuerza protomotríz (PMF) requerida para la síntesis de ATP y de la retroalimentación de electrones de vuelta al sistema. Finalmente, las reacciones dependientes de la luz, como también todas las maquinarias respiratorias, cierran el proceso mediante la enzima ATP Synthase, la cual es responsable de la síntesis de ATP, lo que permite en último término que la energía lumínica se transforme en energía química, se exprese en complejidad bioquímica y sostenga el proceso global celular operando adecuadamente .
Una vez acumulada la energía en forma de ATP entran en juego las reacciones no dependientes de la luz, anteriormente llamadas reacciones de obscuridad, que en este caso están representadas por el ciclo CBB. Este ciclo, activado por las moléculas mencionadas, permite capturar carbono a partir de CO2 y sintetizar glucosa. El ciclo se tratará de forma detallada en otra lectura. La fIG.5 exponede manera detallada la maquinaria fotosintética de bacterias púrpura . En la esquina superior izquierda de la figura se ilustra la invaginacion de la membrana interna y el espacio periplasmático donde tiene lugar el proceso.
Fig 5. Bacterias púrpura: Aparato fotosintético
REACCIONES DEPENDIENTES DE LA LUZ
CAPTURA DE LUZ (LIGHT HARVESTING)
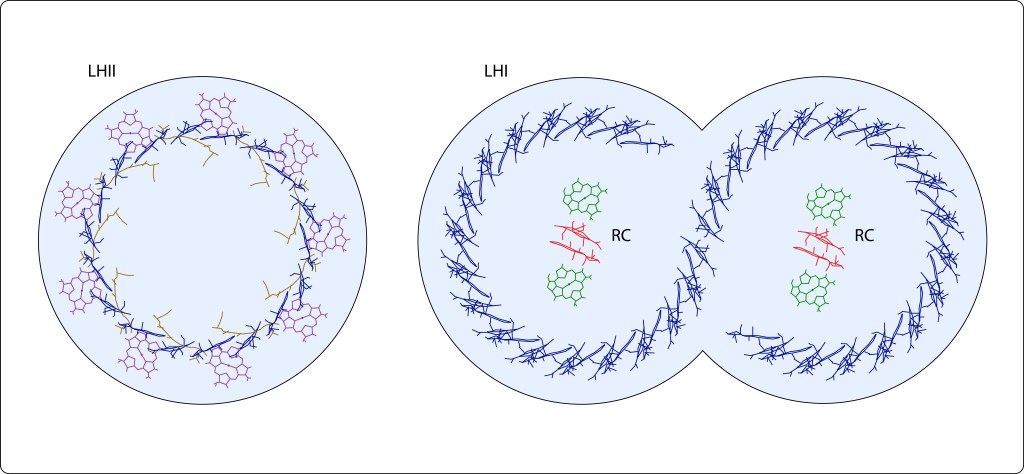
En bacterias púrpura, la energía lumínica exita pigmentos fotosintéticos, carotenos y clorofilas, geométricamente distribuídos al interior de dos tipos de complejos proteicos transmembrana denominados complejos antena LHII y LHI, ver Fig 6.
Fig 6. Vista superior de los complejos antena de Bacterias Púrpura. LHII (izquierda) contiene 9 carotenos (café) en empalizada , 9 Bchla´s paralelas y 18 Bchl´s perpendiculares a la membrana plasmática. LHI dimérico en forma de “S” contiene 36X2 clorofilas dispuestas perpendicularmente a la membrana plasmática. Círculos en azul claro representan las proteinas transmembrana correspondientes a cada monomero.
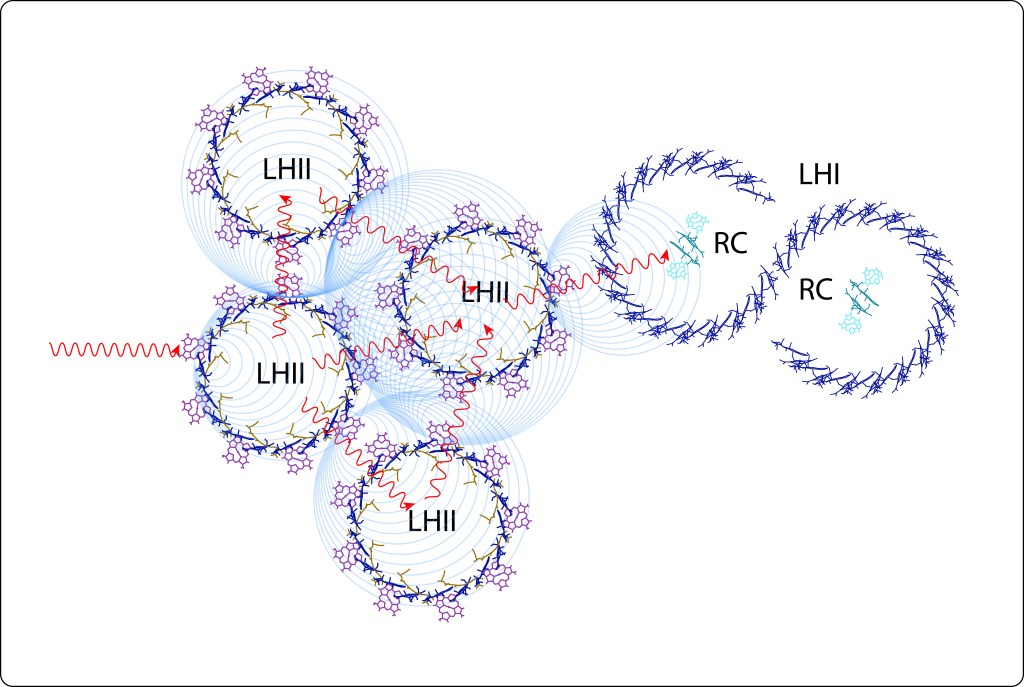
Mediante resonancia no radiativa (Resonancia de Förster) y coherencia cuántica, la energía capturada en LHII es transferida a LHI, para finalmente ser transladada a un par especial de bacterioclorofilas, denominadas Special Pair, localizadas en el Centro de Reacción, RC, ver Fig 7.
Fig 7. Flujo de energía desde los Complejos Antena LH hasta en centro de reacción RC.
La transferencia radiativa implica una disminución gradual de la energía en el tránsito desde LHII hasta LHI.
2. EXITACIÓN DEL PAR ESPECIAL DE CLOROFILAS
El par especial P870 en RC exitado por un valor de energía superior al valor de exitación de su donor en LHI (B875)., ver Fig. 8, lo cual se debe probablemente al efecto combinado de coherencia y superradiancia cuánticas.
Fig 8. Energías y rutas de exitación en los Complejos Antena LH hasta en centro de reacción RC. Transcrito de Proc. Natl. Acad. Sci. USA 95 (1998)
3. FLUJO DE ELECTRONES ETC
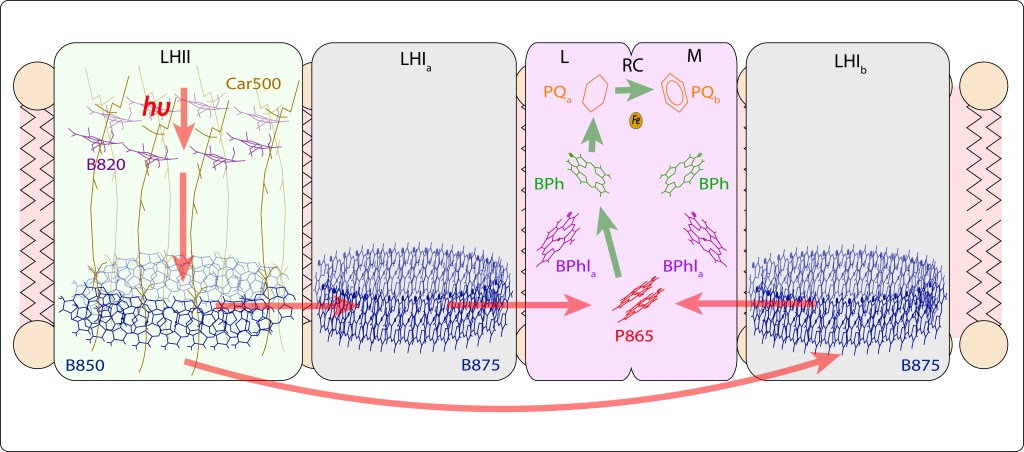
El flujo de energía lumínica, indicado con flechas en rojo en la Fig 8., tiene lugar vía LHII → LHI → P870 en RC, mientras que la cadena de transporte de electrones ETC, flechas en verde, ocurre desde el par especial hasta PQ b en el centro de reacción RC. Ver Fig.9.
Fig 9. Flujo de energía en los complejos antena de Bacterias Púrpura. Se incluye la transferencia de LHI a RC, y la cadena de transporte en RC, tema amplado más adelante. Las subunidades LHIa y LHIb en la imagen corresponden a un único complejo LHI en “S” , ver también Fig 3., rodeando el centro de reacción RC . Note que el complejo proteico de RC está formado por un dímeero con subunidades L, M.
NIVELES DE ENERGÍA
Los pigmentos fortosintéticos presentan, dependiendo de la energía incidente, dos niveles de exitación: 1° y 2° estados en singlete, ver Fig 10.
Fig 10. Niveles de energía en la molécula de clorofila. Dependiendo de la luz incidente, el estado de exitación puede ser 2° singlete (banda azul), o 1° singlete (banda roja). Fuente : Biochemistry & Molecular Biology of Plants.2015, John Willey & Sons.
Electrones en 2° singlete se relajan rápidamente de manera expontánea a 1° singlete por conversión interna (IC) y liberación de calor, mientras que los de 1° singlete (en el caso del par especial de clorofilas) pueden tener dos destinos:
Retorno a GND por fluorescencia
Liberación desde el par especial vía REDOX (reducción de BPha como primer aceptor de electrones) , iniciandose con este evento la cadena de transporte de electrones ETC. La ruta tomada dependerá de los requerimientos fisiológicos de ATP. Alta demanda favorecerá ETC, caso contrario la molécula retornará a GND.
Ante demanda de ATP, el electrón cedido por el par especial inicia una serie de reacciones REDOX en cadena, ver Fig 10.
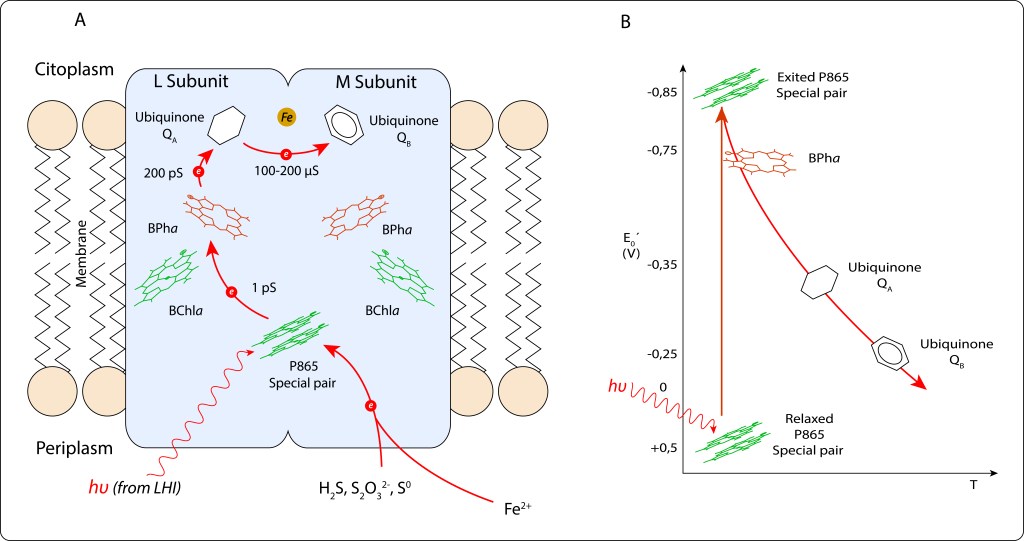
P865 → BPha → QA → QB.
Cofatores adicionales, BChla y Fe2+ , indicados en la figura 11., no participan directamente en la cadena de transporte, su función es estabilizar la carga local para facilitar la unidireccionalidad de la cadena de reacciones.
Fig 11. Cadena de transporte de electrones en RC. A. Pigmentos y moléculas involucradas en el transporte. B. Niveles de energía correspondientes a cada etapa.
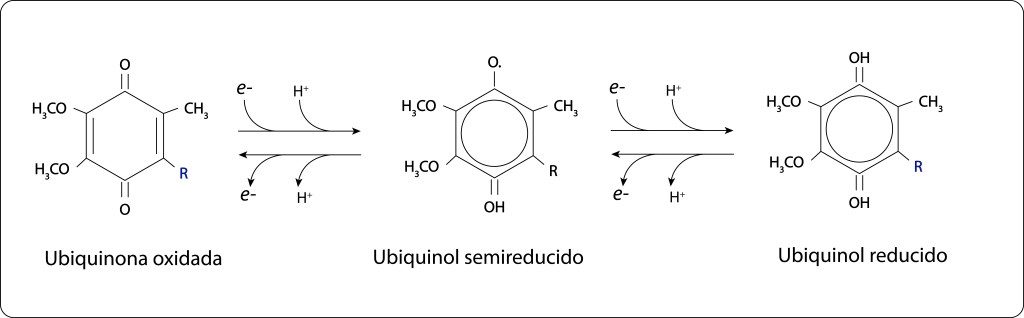
El aceptor final en RC, Q b , requiere dos electrones para reducirse completamente, ver Fig 12., de modo que el par especial debe ceder secuencialmente dos electrones , lo que implica captura de dos fotones por parte de los complejos antena y consecuentemente utilización de los dos electrones que proceden del citocromo C en una ruta cíclica.
Fig 12. Reducción de Ubiquinona a Ubiquinol, pasando por Ubiquinol semireducido.
La plastoquinona completamente reducida migra al pool de quinonas y mediante la adquisición de 2 cationes H+ procedentes del citoplasma se convierte rapidamente en Plastoquinol . El confinamiento de plastoquinonas y plastoquinol al interior de la membrana citoplasmática interna es posible gracias a su alta hidrofobicidad molecular, de manera que los fosfatos de la bicapa fosfolipídica actuan como barrera de retención manteniendo las quinonas y quinoles en el pool a disposición del citocromo Bc1.
4. FUERZA PROTOMOTRIZ PMF
Ubiquinol se convierte entonces en el portador de energía a ser utilizado por el citocromo bc1 para la generación del gradiente protónico requerido para la síntesis posterior de ATP.
El complejo citocromo bc1 , al igual que b6f en cianobaterias y en mitocondrias de células eucariotas, se caracteriza por contener moléculas Heme como cofactores responsables del transporte de electrones.
En bc1, tres Heme participan en el transporte de electrones, dos en Cyt b y una en Cyt c1; lo que contrasta con el centro de reacción RC, donde los cofactores responsables del transporte corresponden a clorofilas, phaeophytinas y quinonas.
En Heme, un átomo de Hierro Fe2+ es quelatado de manera similar a la quelatación de magnesio Mg2+ en las clorofilas y bacterioclorofilas, ver Fig 13.
Fig 13. Esquema comparativo de las moléculas de Bacterioclorofila a y Heme.
Mientras que en RC los electrones cedidos por el par especial de clorofilas corresponden a electrones π de la cadena resonante de enlaces conjugados, en heme los electrones provienen del ión ferroso Fe2+ al pasar a estado férrico Fe3+.
Heme y clorofila comparten una misma ruta de síntesis metabólica, partiendo del ácido δ-aminolevulínico (ALA) hasta el subproducto protoporfirina IX. De ahí en adelante los agentes quelatantes y demás diferenciadores definen el producto final.
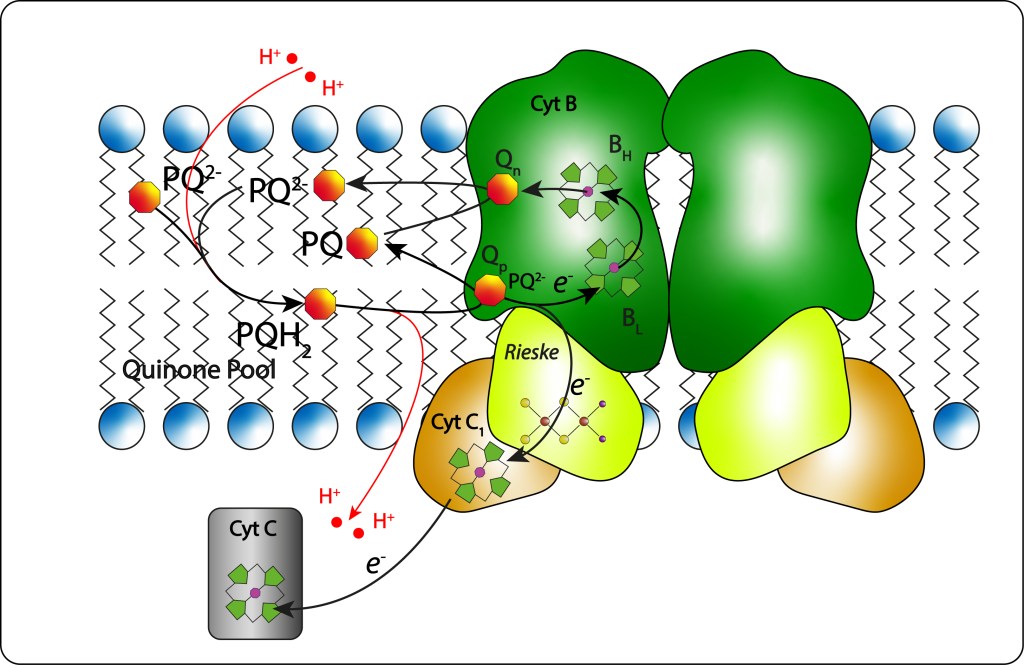
El Ubiquinol (QH2) exportado desde el centro de reacción al pool de quinonas es redireccionado al sitio Qp del CytB, ver Fig 14, donde es totalmente oxidado.
Fig 14. Cytochrome BC1. Función y diagrama general
Un primer electrón es capturado por el cofactor Fe2S2 del complejo Rieske, ver Fig 15., de allí pasa al citocromo C1 y luego al Cyt C2 soluble que lo lleva de nuevo al centro de reacción RC para reducir el par especial de clorofilas, pudiendo reiniciarse así un nuevo ciclo de transporte de electrones.
Fig 15. Complejo Rieske. Uno de los dos hierros es coordinado por dos residuos His definiendo su alto potencial Redox.
El segundo electron sigue una ruta diferente: Heme Bl (bajo potencial) -> Heme BH (alto potencial) → quinona Q (localizada en un segundo sitio de CitB: Qn) reduciendola parcialmente (Q → Q–) , de manera que la reducción completa de Q implica la reducción de dos ubiquinoles.
La figura 16 ilustra los potenciales Redox en bacterias púrpura y bacterias verdes del azufre (para fines comparativos) a lo largo de la cadena de transporte de electrones, incluyendo las etapas que tienen lugar en RC y las etapas en Cyt bc1. Note que en bacterias púrpura la reducción de NAD+ de manera directa por la vía de fotosíntesis, por su alto valor de potencial.
Como se puede observar, de manera global los potenciales Redox de bacterias púrpura son menores comparados con los potenciales de bacterias verdes del azufre.
Esto define para el sistema de bacterias púrpura un poder oxidante fuerte, reductor débil, mientras que GSB se define un poder oidante débil, reductor fuerte.
la importancia de esta diferencia en poder oxidoreductor entre uno y otro grupo biológico se entenderá más adelante, al abordar las cyanobacterias, un grupo biológico que por endosimbiosis bioquímica (término personal propuesto en estas lecturas) integró los dos fotosistemas en una sola maquinaria fuertemente oxidante, fuertemente reductora, capaz de oxidar H2O y capaz de reducir NAD+ de manera autónoma, lo que en su momento evolutivo defino un nuevo curso para la vida en el planeta.
Figura 16. Potenciales redox en bacterias purpura y verdes del azufre
5. SÍNTESIS DE ATP
Como resultado de la oxidación del Ubiquinol, dos hidrogeniones H+ son exportados al periplasma contribuyendo a generar el gradiente PMF requerido para la síntesis de ATP.
El citocromo BC1 actua entonces como una bomba de protones, desde el citoplasma al periplasma, y el PMF generado actúa como habilitante de la ATP Synthase, último complejo de la cadena de reacciónes dependientes de la luz.
El diferencial de iones H+ entre periplasma y citoplasma, PFM, es utilizado por ATP synthase para la síntesis de ATP.
Si el Citocromo BC1 es una bomba de protones, ATP synthase es una verdadera ensambladora rotativa de tres tiempos, admisión, compresión y expulsión, con un rotor dotado de tres sitios activos separados 120° entre sí. La rotación es habilitada gracias al flujo de protones procendentes de PMF a travez de la ATP Synthase.
El primer sitio activo, o sitio de admisión, recibe ADP y radicales fosfato. El sitio de compresión inserta mediante ataque nucleofílico el fosfato en la molécula de ADP para generar ATP, y finalmente en el sitio de expulsión el ATP se libera al citoplasma quedando a disposición de los procesos bioquímicos celulares, y en particular a las cadenas fotosintéticas no dependientes de la luz, que en el caso de las bacterias púrpura, corresponden al ciclo de Calvin-Benson.
Esta enzima no es exclusiva de las bacterias púrpura, y está presente prácticamente en todos los organismos vivientes, desde arqueas hasta plantas superiores y animales, y siempre su función es la síntesis de ATP, utilizando el gradiente electroquímico de protones H+ , aunque en algunos casos también utiliza el gradiente de iones Na+ (Fuerza Sodio-motriz) , y raramente K+.
6. SÍNTESIS DE NADPH – NADH
Como se ilustró en la Fig.16., el centro de reacción P870 característico de bacterias púrpura NO es lo suficientemente reductor para reducir directamente NAD⁺, de modo que se debe contar con una maquinaria adicional para producirla.
Existen varias rutas reductoras principales:
NADPH vía electrones provenientes de Cyt bc1:
Por esta ruta, parte de los electrones provenientes de Cyt bc1 via complejo de Rieske no retroalimentan RC sino que son redireccionados a Fdox para su reducción:
Fdox +e– → Fdred
A continuación, la enzima FNR (ferrodoxina-NADP+ reductasa), cataliza la reacción
Fdored + NADP+ + H+ → Fdox + NADPH
NADH víaElectrones provenientes de donores orgánicos (Succinato, lactato):
En esta ruta, dado el bajo poder reductor de la maquinaria fotosintética y alto potencial Redox del par NAD+/NADH, se habilita el complejo II (Complejo SDH) de la máquinaria respiratoria (Succinato deshidrogenasa), permitiendo que succinato (o lactato) reduzcan quinonas a ser redireccionadas no hacia Cyt bc1 sino hacia el complejo I de la cadena respiratoria. Utilizando ATP-asa (ATP-Sinthase inversa) se incrementa el gradiente protónico en el espacio periplasmático lo que genera un flujo protónico inverso que a su vez permite que los electrones fluyan “Up Stream” en la cadena de ferrodoxinas hasta finalmente reducir NAD+. Ver Fig.17. Este proceso ocurre en obscuridad, de manera que el aparato fotosintético (módulos en gris en la figura) permanecen inactivos. Con propositos comparativos se anexa como Fig.18 el complejo fotosintético previamente presentado en la Fig 4. Vale la pena resaltar la función de “Switch” o conmutador de energia de l complejo II, el cual no solo alimenta alternativamente el Complejo I en la síntesis de NADH o el pool de quinonas en el proceso fotosintético sino que también invierte el flujo protónico, convirtiendo ATP-Synthase en ATP-Asa, forzando a un flujo protónico de citoplasma a pariplasma, y tambien invirtiendo este flujo en el complejo I, lo que finalmente lleva a sintetizar (y no consumir) NADH.
Figura 17. Síntesis de NADH en bacterias púrpura fotosintéticas via complejos de la cadena respiratoria.
Figura 18. Diagrama de flujo reacciones fotosintéticas dependientes de la luz en bacterias púrpura
NADPH vía transhidrogenasas.
Las transhidrogenasas son enzimas que transfieren hidruros (H⁻) entre NADH y NADP⁺ para producir NADPH, siendo en la práctica un intercambio de poder reductor entre estos dos cofactores:
NADH + NADP+ ↔ NAD+ + NADPH
Esta ruta es particularmente útil cuando:
Hay exceso de NADH (por respiración o metabolismo de sustratos orgánicos).
Hay demanda de NADPH para biosíntesis (fijación de CO₂ cia CBB, síntesis de lípidos, etc).
REACCIONES NO DEPENDIENTES DE LA LUZ
En bacterias púrpura, al igual que en cyanobacterias y plantas superiores, el ciclo de Calvin-Benson es el receptor de facto del ATP producido en la fotosíntesis. Este ciclo forma parte integral de la maquinaria fotosintética, y corresponde a lo que se denomina reacciones no dependientes de la luz.
El ciclo de Calvin-Benson merece una lectura aparte, luego no se detalla en el presente blog.






 , corresponden a los siguientes eigenspinors:
, corresponden a los siguientes eigenspinors: