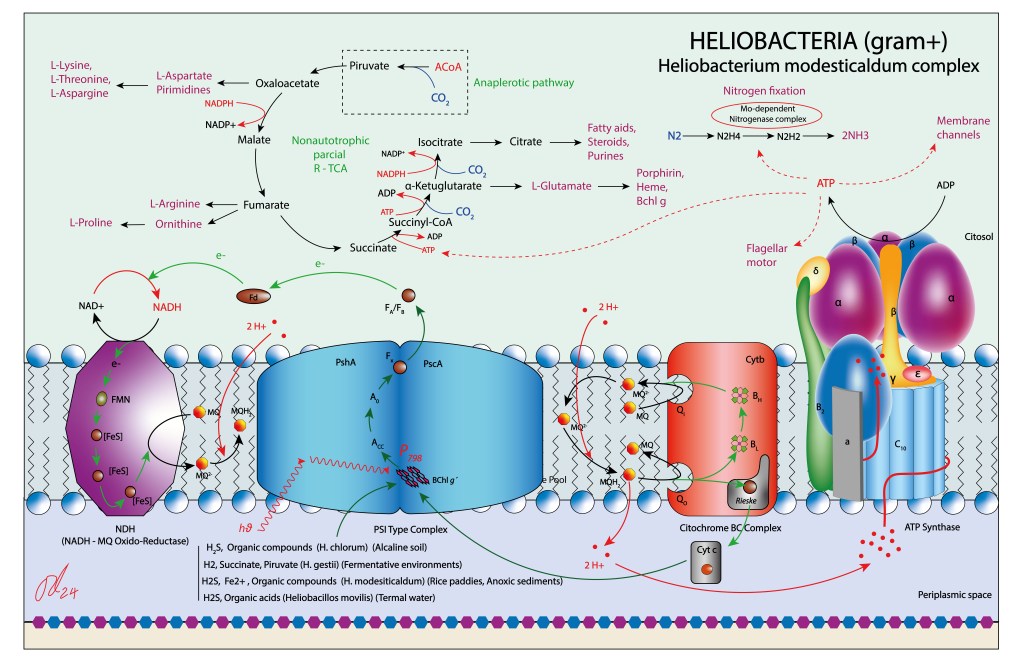
La clase Heliobacterias (Orden Heliobacteriales, Phyllum Firmicutes), corresponde a un grupo muy especializado de bacterias fotoheterótrofas Gram+.
Descrito por primera vez en 1983 por Ghest y colabordores [3], a la fecha tiene definidos dos clados, cuatro géneros (Heliobacterium, Heliobacillus, Heliophilum y Heliorestis) y diez especies.
Las especies son mayormente terrestres, raramente acuáticas, anaerobias estrictas, prosperando en suelos anóxicos, muchas veces asociadas en relaciones mutualistas con cultivos de arroz, y al igual que las especies heterótrofas del grupo, presentan endosporulación. Algunas especies (Heliobacillus movilis, Heliobacterium modesticalium) han sido reportadas en suelos proximos a fuentes termales. H. modesticalium, hallada Islandia, ha sido una de las especies mejor estudiadas.
Considerado a nivel bioquímico un grupo minimalista, exhibe el aparato bioquímico de captura de energía mas simple dentro de todos los grupos bacterianos fototrófos y fotosintéticos reportados a la fecha. Su hallazgo ha aportando significativamente al entendimiento de la evolución de la fotosíntesis, se considera un grupo muy primitivo, con razgos bioquímicos ancestrales conservados.
Las Heliobacterias son incapaces de fijar carbono a partir de CO2, por lo que se considera un grupo bacteriano fotoheterótrofo, no fotosintético. Dependiendo de las condiciones ambientales, utilizan distintos compuestos como fuentes de electrones: H2S y compuestos orgánicos en suelos alcalinos, Fe2+, H2, succinatos y piruvatos en ambientes reductores y ácidos orgánicos en aguas termales.
CAPTURA DE ENERGÍA
A diferencia de los demás grupos bacterianos fotótrofos, las Heliobacterias no presentan complejos antena exclusivos a la función de captura de energía lumínica.
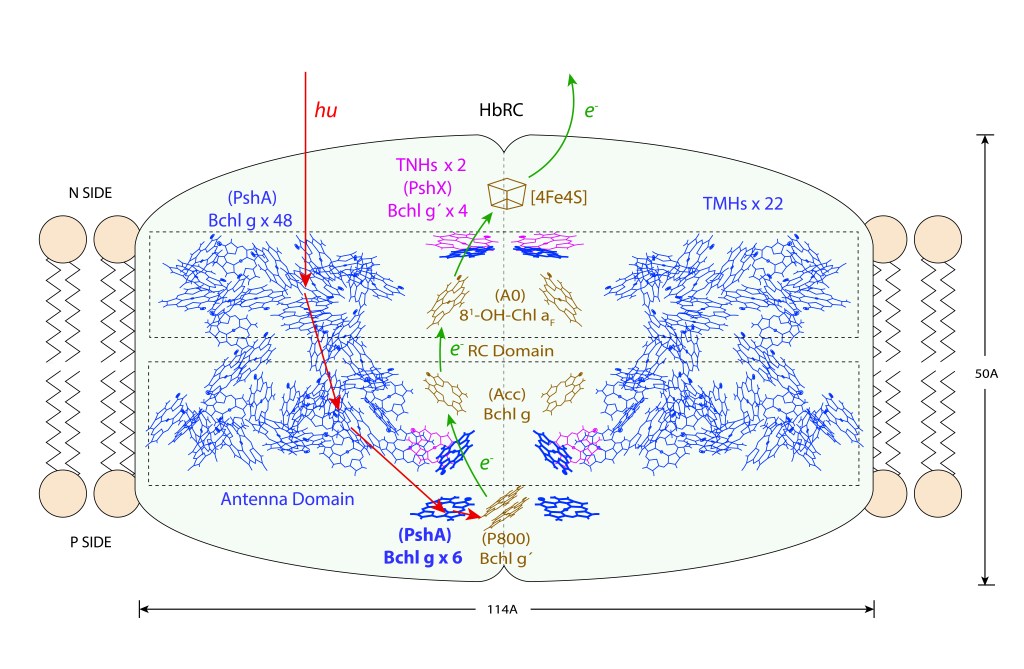
Su centro de reacción, HbRC, integra en un solo complejo las funciones de captura de energía y tránsito ETC, con dos dominios claramente diferenciables para cada una de las funciones: Dominio antena y dominio RC.
ESTRUCTURA PROTEÍNICA
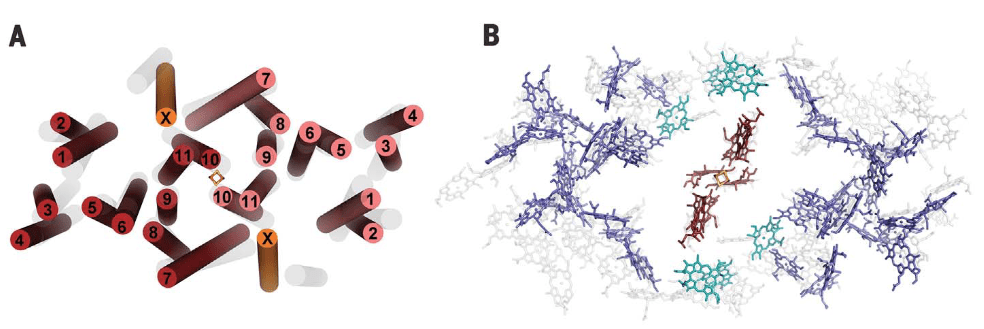
El complejo HbRC corresponde a un homodimero con simetría C2, 24 hélices transmembrana (TMHs) de las cuales 22 están asociadas al homodímero PshA y dos recientemente descritas a una nueva subunidad PshX, ver Fig.1 A. La distribución de pigmentos se describe en la Fig.1 B.
Figura 1. Arreglo de hélices Transmembrana (TMHs y pigmentos) en HbRC. Vista superior. Fuente: Gisriel et.al., 2017. En las imagenes, el autor superpone en gris las hélices (Fig A) y pigmentos (FigB) correspondientes a TMHs de PSI de plantas superiores.
DOMINIO ANTENA
El core del Dominio Antena corresponde a 54 bacterioclorofilas Bchl g (pigmentos de color azul) distribuidas en dos capas, ver Fig 2. De las 54 Bchl g, seis (azul intenso) son proximales al dominio RC, siendo probablemente las unidades responsables de la exitación del par especial de clorofilas.
Es importante resaltar que Gchl g , un isómero de Chl a, solo está presente en este grupo bacteriano.
En adición a las 54 Bchlg, cuatro Bchl g´ adicionales estan coordinadas por las subunidades Pshx (pigmentos magenta) igualmente proximales a RC. Bchl g´es un esteroisómero de Bchl g.
Figura 2. Heliobacterias: Estructura y dinámica general del complejo HbRC. Nota: La distribución y número de pigmentos en PshA es solo ilustrativo. para información mas rigurosa consultar Gisriel et.al., 2017.
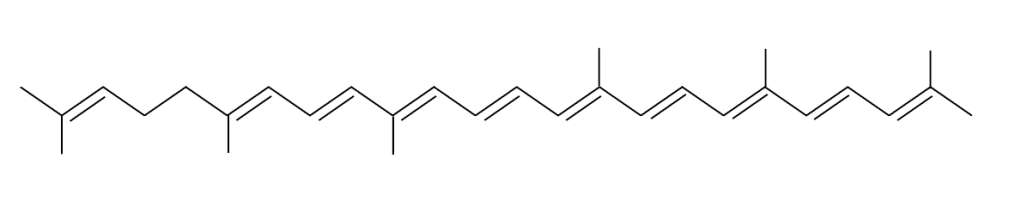
En adición a los pigmentos mencionados, dos moléculas de naturaleza lipídica y dos carotenos 4,4´ Diaponeurosporeno, uno por sumbunidad mioonodimérica, forman parte del Dominio Antena, ver Fig.3.
Figura 3. Estructura molecular C30 4,4´ diaponeurosporeno
DOMINIO RC
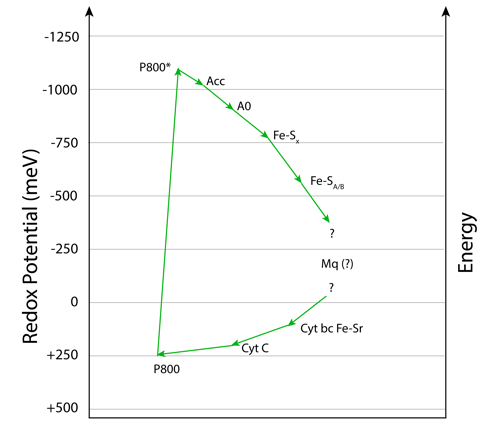
La ruta de transferencia electrónica (ETC) en Heliobacterias es similar a la de bacterias verdes del azufre, si bien la frecuencia de exitación es mucho más baja, ubicandose en el infrarojo cercano, alrededor de 798-800 nm. ver Fig.4.
En heliobacterias, P800 corresponde a Bchl g´, Acc a Bchl g y A0 a la clorofila 81-OH-Bchl aF.
La energía es finalmente entregada a una Fx del tipo [4Fe 4S] para luego reducir NAD+ a su forma NADH, mediante un intermediario FeS A/B.
Figura 4. Heliobacterias: Ruta de transferencia electrónica.
POOL DE QUINONAS
El pool de quinonas está representado por menaquinonas. En ausencia de un complejo II que pueda aportar directamente quinonas reducidas al pool, la enzina NDH puede estar soportando este rol, con NADH como fuente aportante de electrones.
CITOCROMO BC
Similar en función a los citocromos BC1 y B6f, realizan en este caso un flujo cíclico, con electrones transportados por un Cyt C soluble hacia el fotosistema, contribuyendo al aporte de electrones.
METABOLISMO REDUCTOR
Como ruta reductora, las Heliobacterias disponen de un ciclo parcial r-TCA . Dada la la ausencia de genes codificantes de ATP-Citrato Lyasa, el piruvato requerido por el ciclo debe ser obtenido por rutas anapleróticas, cedido por las plantas hospederas o a partir del sustrato.
Al igual que en GSB, los Ciclos CBB y Biciclo 3-HP están ausentes. En general las rutas bioquímicas del carbono aparecen muy simplificadas en Heliobacterias. En contraste, las rutas fijadoras de nitrógeno (Mo dependientes) son muy robustas. En este sentido podrían desempeñar en gramineas un soporte nitrificante similar al de Rhizobium, Bradyrhizobium y Sinorhizobium en leguminosas, si bién nunca conformando nódulos radiculares.
Puesto que son un grupo Gram+ carente de membrana interna, el espacio periplasmático desempeña la función del lúmen thylakoidal de cyanobacterias y del espacio intermembrana de bacterias púrpura, lo que les permite confinar el H+ requerido para la síntesis de ATP.
Como se mencionó anteriormente, la enzima transmembrana NDH (NADH-MQ Oxidoreductasa) forma parte de la bioquímica del complejo jugando un rol doble, transporte H+ hacia el espacio periplasmático y reducción de menaquinonas, lo que debe incrementar de manera significante la producción de ATP, ver Fig 5.
Figura 5 . Aparato fototrófico propio de Heliobacterias.
BIBLIOGRAFÍA RECOMENDADA
[1] Asao M, Madigan MT. Taxonomy, phylogeny, and ecology of the heliobacteria. Photosynth Res. 2010 Jun;104(2-3):103-11. doi: 10.1007/s11120-009-9516-1. Epub 2010 Jan 22. PMID: 20094790.
[2] Gisriel. C., Sarrou, I., Ferlez, B., Golbeck, J, J.H., Redding, K.E., and Fromme, R. (2017) Structure of a symmetric photosynthetic reaction center–photosystem. Science. 357:1021-1025
[3] Gest H. Discovery of the heliobacteria. Photosynth Res. 1994 Jul;41(1):17-21. doi: 10.1007/BF02184140. PMID: 24310008.
[4] Sattley WM, Blankenship RE. Insights into heliobacterial photosynthesis and physiology from the genome of Heliobacterium modesticaldum. Photosynth Res. 2010 Jun;104(2-3):113-22. doi: 10.1007/s11120-010-9529-9. Epub 2010 Feb 4. PMID: 20130998.
[5]Sattley WM, Madigan MT, Swingley WD, Cheung PC, Clocksin KM, Conrad AL Dejesa LC, Honchak BM, Jung DO, Karbach LE, Kurdoglu A, Lahiri S, Mastrian SD, Page LE, Taylor HL, Wang ZT, Raymond J, Chen M, Blankenship RE, Touchman JW 2008. The Genome of Heliobacterium modesticaldum, a Phototrophic Representative of the Firmicutes Containing the Simplest Photosynthetic Apparatus. J Bacteriol 190:. https://doi.org/10.1128/jb.00299-08
La convergencia evolutiva de los fotosistemas PSI y PSII ha sido sin lugar a dudas la innovación bioquimica de mayor impacto en la historia de vida en nuestro planeta.
Esta trascendental propuesta, desarrollada hace algo más de 2,500 millones de años por las cyanobacterias y luego transmitida por endosimbiosis seriada hacia algas y plantas superiores, permitió extraer del agua los electrones requeridos para mantener en funcionamiento la maquinaria fotosintética, llevandola a un nivel de eficiencia que terminó redireccionando el curso evolutivo de la vida.
El precio de dicha innovación fué muy alto.
El oxígeno liberado durante el proceso de oxidación del agua se convirtió en una amenaza para la vida, e incluso en una amenaza para la misma maquinaria que lo generó, lo que implicó involucrar una serie de rutas de protección de la maquinaria fotosintética, ver Fig 1.
Bajo condiciones ideales, y con un aparato fotosintético produciendo justo lo requerido, el oxígeno residual no es realmente una amenaza.
Pero variaciones en la disponibilidad de luz, cambios en PH, temperatura y humedad, restricciones en el acceso a CO2 , etc, implican cambios en los balances de las reacciones de oxidoreducción.
En estos casos, la oferta de electrones generados puede llegar a superar la capacidad de consumo por parte de la misma, lo que resulta en una cadena de trasporte de electrones ETC altamente reductora.
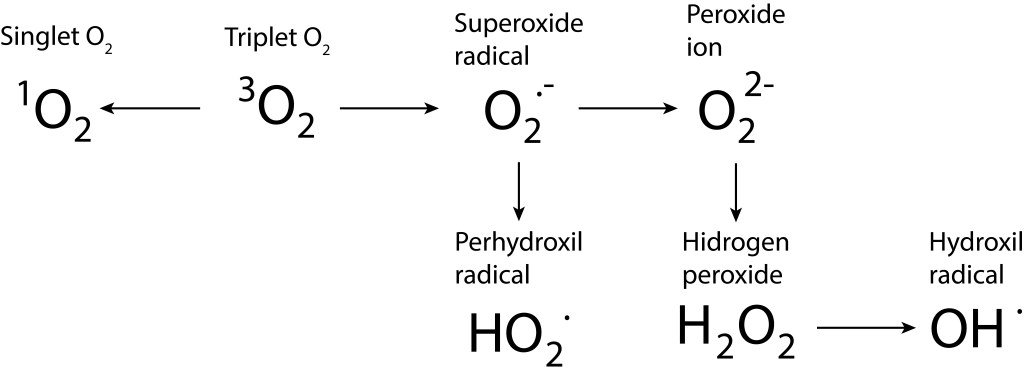
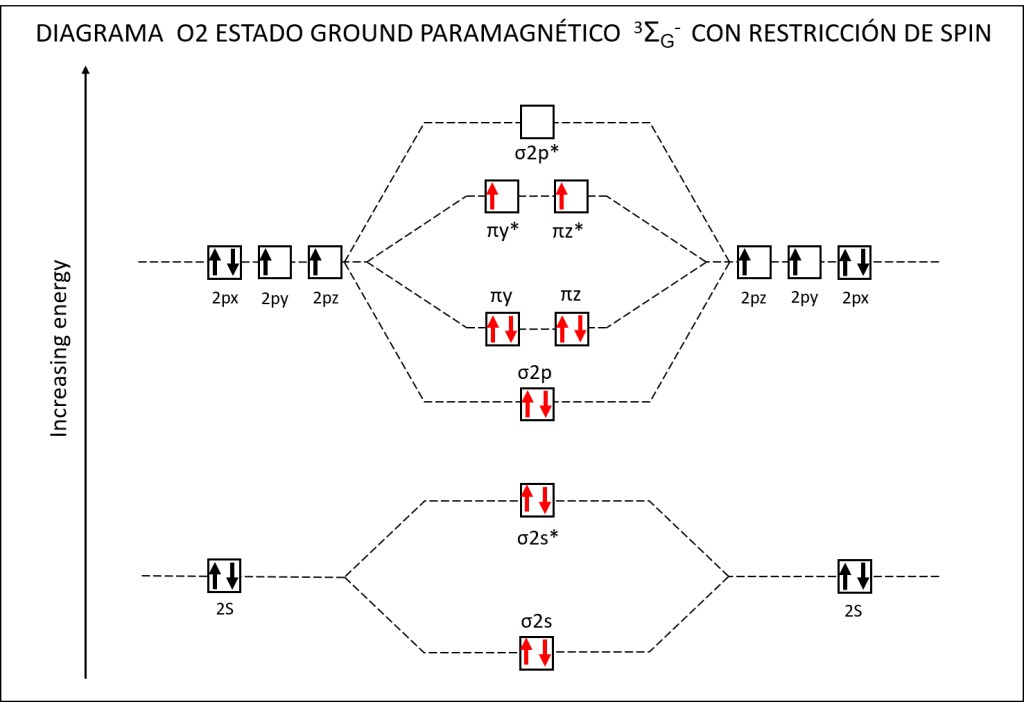
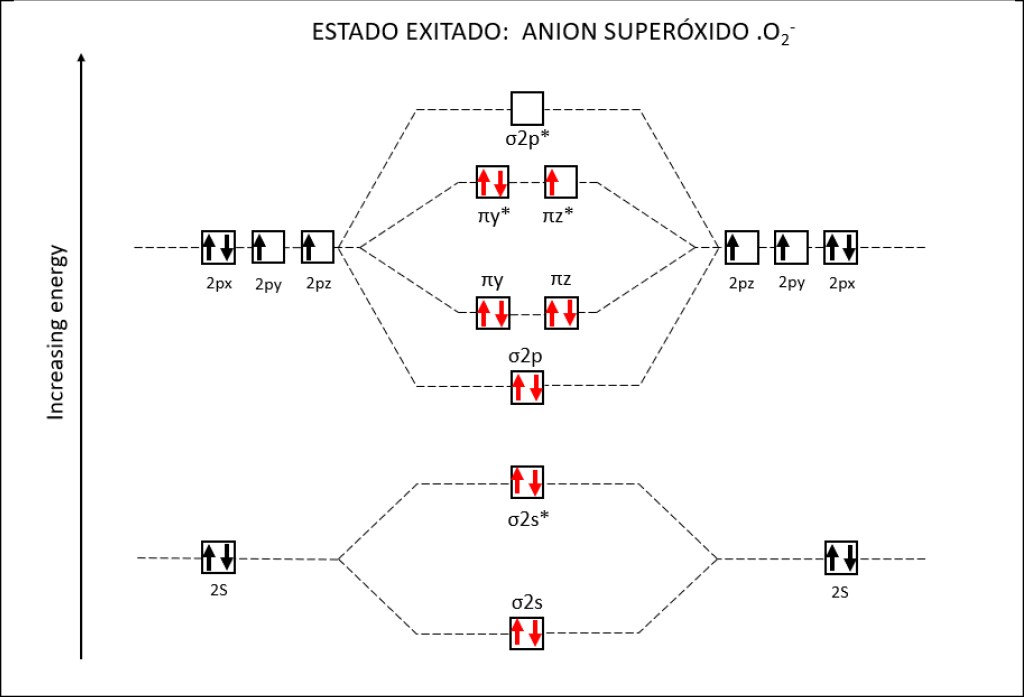
Bajo estas condiciónes, la probabilidad de generarse especies reactivas de oxígeno ROS, es muy alta, exponiendo a PSI y PSII a daños severos. La síntesis de especies reactivas del oxígeno y los orbitales moleculares correspondientes a las mismas se resumen en las Fig 1 y Fig 2.
Figura 1. Rutas de formación de especies reactivas de oxígeno.
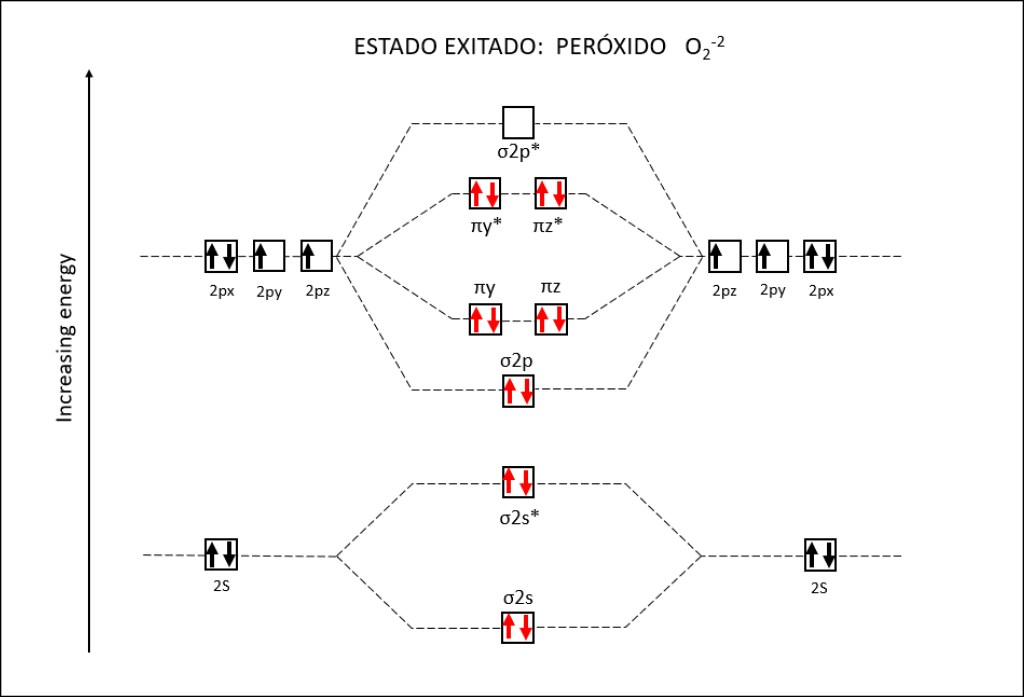
Figura 2 : Orbitales moleculares correspondientes al estado ground y a los estados exitados superóxido y peróxido de la molécula de oxígeno.
La generación de radicales hydroxil se dá por dos vías principales:
Por acción de la luz UV , que disocia H2O2 a su forma 2 OH.
En presencia de Fe(II), en la reacción de Fenton:
Fe2+ + H2O2 → Fe3+ + OH. + OH–
Los radicales hydroxil son extremadamente reactivos, responsables del estrés oxidativo y causantes del daño no solo de los fotosistemas sino en general del entorno celular.
Mientras PSII tiene una capacidad de respuesta regenerativa relativamente alta ante la acción de especies reactivas, PSI es mucho más sensible a daños definitivos, exponiendo a la maquinaria global a colapsar en su función.
PROTECCIÓN ROS
La generación de especies reactivas de oxígeno hace necesario contar con rutas bioquimicas de control que permitan mantener el entorno fotosintético en estado oxidado, ver Fig 3.
Figura 3. Rutas NPQ y protección ROS presentes en la maquinaria fotosintética de cyanobacterias.
Las rutas de control ROS pueden clasificarse en dos tipos:
RUTAS PREVENTIVAS: Actúan sobre entornos reductores redireccionando el flujo de electrones hacia pool de quinonas para luego extraerlos por agentes oxidantes específicos.
RUTAS CORRECTIVAS: Actúan reduciendo las especies ROS generadas.
RUTAS PREVENTIVAS ROS
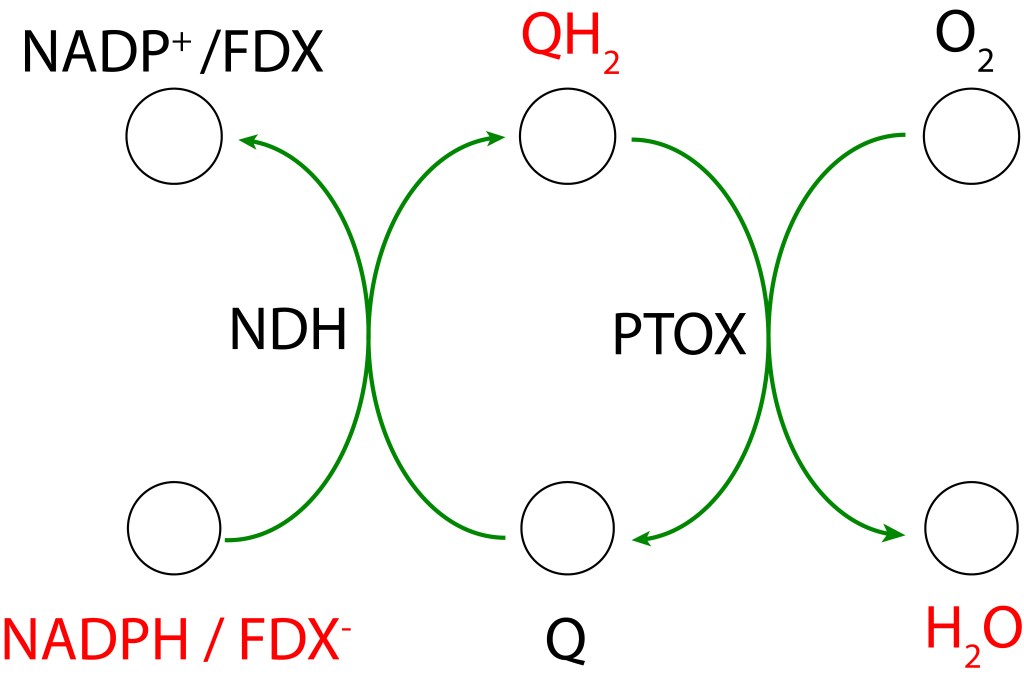
RUTA PTOX MEDIADA POR NDH-1
PTOX Deriva su nombre de la enzyma “Plastid Terminal Oxidase”, o“Plastoquinol Terminal Oxidase“, la cual reside adyacente a la membrana thylakoide.
Esta enzima actúa como válvula de seguridad, extrayendo electrones del pool de quinonas mediante la reacción
PQH2 + O2 → 2 PQ + 2 H2O
PQ es reducido a su forma PQH2 mediante un flujo cíclico de electrones (Non photochemical electron cycle) provenientes de FDX– siguendo la ruta
FDX– → NDH-1 → PQ → PQH2
NDH-1 corresponde al complejo proteico transmembrana NADPH dehydrogenasa presente en cyanobacterias, es responsable de la oxidación de FDX– y de la correspondiente reducción de PQ.
La transferencia de electrones en la ruta PTOX se conoce como “Clororespiración” y se puede resumir asi:
La ruta se activa, entre otras causas, cuando la disponibilidad de CO2 citoplasmatico no es lo suficientemente alta como para atender los requerimientos del ciclo de Calvin Benson. Bajo esta condición, la caída en concentración de CO2 desacelera el ritmo del ciclo CBB generando un represamiento de las especies NADPH y FDX– , lo que promueve un entorno reductor favorable a la formación de especies ROS, particularmente por FDX– , cuyo alto represamiento puede terminar transfiriendo electrones a O2 generado las especies ROS mencionadas.
Para prevenir esta condición, las rutas PTOX reenrutan los electrones hacia el pool de quinonas Se ha reportado que la acción de PTOX es responsable hasta del 30% del drenaje de electrones en el tráfico ETC (Zolotereva et.al. 2022).
La acción colaborativa PTOX/NDH-1 actúa sobre FDX– “aguas abajo” de PSI y Cyt b6f permitiendo que bajo condiciones de stress, la síntesis de ATP se mantenga intacta, lo que hace de este esquema de protección un mecanismo altamente eficiente para la función general fotosintética.
NDH-1 participa en el bombeo de protones al lumen durante la transferencia electrónica de manera equivalente al complejo I mitocondrial y al NDH presente en plantas y algas verdes, contribuyendo a incrementar la fuerza protomotriz ΔpH , lo que aumenta la eficiencia energética del sistema.
RUTA PTOX MEDIADA POR PGRL1-PGR5
La proteína transmembrana PGRL1 (Proton Gradient Regulation Like 1) es una proteína reguladora transmembrana de bajo peso molecular, presente en cyanobacterias, plantas y algas verdes.
PGRL1 actúa en conjunto con la proteina PGR5 (Proton Gradient Regulation 5), conformando un heterodímero con función equivalente a la de NDH-1.
La velocidad de respuesta del heterodímero ante variaciones en concentración FDX– es mucho mayor que la de NDH-1, lo que permite que las dos proteínas actúen de manera complementaria para el funcionamiento óptimo de PTOX. Adicionalmente, mientras PGRL1 opera en condiciones de alta intensidad lumínica, desempeñando una función fotoprotectora, NDH-1 se activa en condiciones de baja luminosidad o bajo stress prolongado.
En resumen, NDH-1 y PGRL1 actúan rediraccionando“de manera preventiva” el flujo de electrones de FDX– hacia el pool de quinonas generando un flujo cíclico electrónico no fotoquímico que contribuye a mantener en estado oxidado el entorno PSI , minimizando por esta vía la generación de especies ROS. Ambas rutas habilitan la función de drenaje PTOX, con H2O como depositario final de electrones y oxígeno provenientes del sistema.
RUTAS CORRECTIVAS ROS
Actúan reduciendo H2O2 a H2O mediante peroxidasas específicas, controlando los niveles ROS en citosol y membrana thylakoide.
Previa la activación de las rutas de control, radicales superóxido O.−2 deben ser convertidos en H2O2 y O2 mediante la acción de Superóxido dismutasas, SOD, en la siguiente reacción simplificada:
O.−2 + O.−2 + 2H+ → O2 + H2O2
SOD corresponde a una familia de metaloenzimas antioxidantes cruciales a la salud celular presentes en archaeas, bacterias y eukariotas que, dependiendo del metal o metales coordinados, se clasifican en los siguientes grupos:
FeSOD : Presente en citosol
MnSOD: Asociada en cyanobacterias y plantas a membrada thylakoide en vecindad a PSII
Cu/ZnSOD: Presente en citosol
NiSOD: presente en cytosol. (Exclusivo de prokariotas)
La reacción comprende dos etapas:
Etapa de reducción del metal coordinado, oxidando O.−2 a O2 :
M(n+1)+−SOD + O.−2 → Mn+−SOD + O2
Etapa de reoxidación del metal coordinado, reduciendo O−2 a H2O2 :
Mn+−SOD + O−2 + 2H+→ M(n+1)+−SOD + H2O2
El estado de oxidación del catión metálico evoluciona entre n+1 → n → n+1:
(Cu2+ → Cu+ → Cu2+)
(Mn3+ → Mn2+ → Mn3+)
(Fe3+ → Fe2+ → Fe3+)
(Ni3+ → Ni2+ → Ni3+)
Una vez convertido O−2 en H2O2 por SOD, tres rutas contribuyen al control ROS mediante la reducción de H2O2 a H2O, teniendo como factor común la cesión inicial de electrones a partir de NADPH.
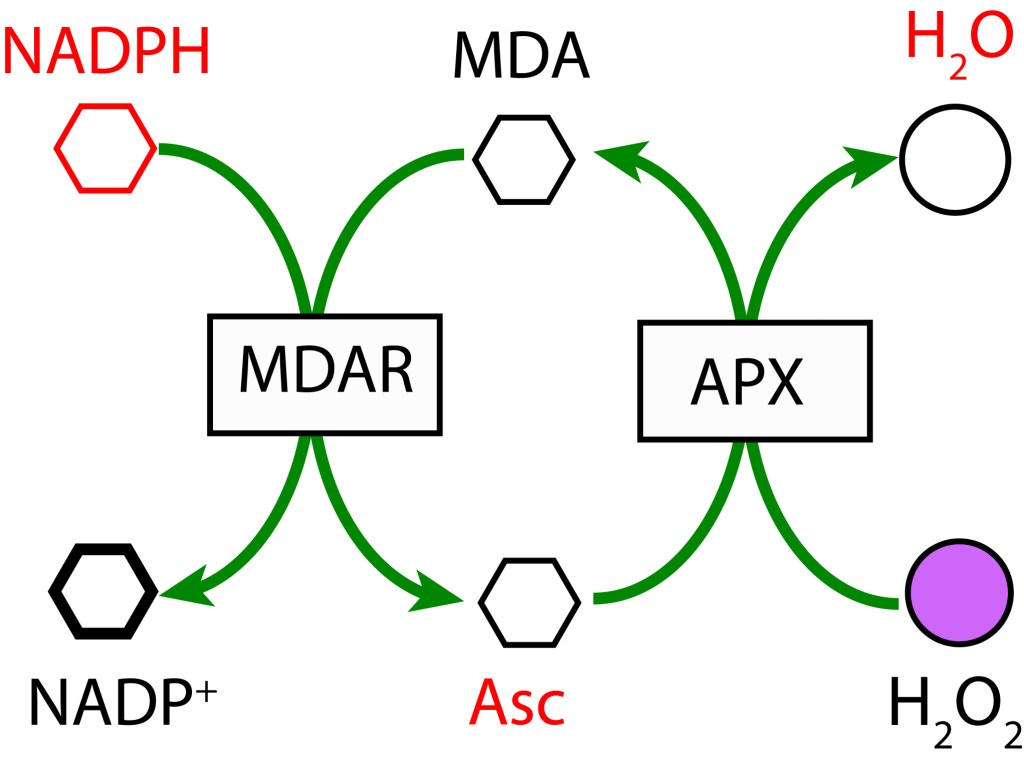
REACCIÓN DE MEHLER (CICLO AGUA-AGUA)
La reacción de Mehler, conocida también como ciclo agua-agua (WWC), está presente en todos los organismos con fotosíntesis oxigénica y presencia de fotosistema PSI.
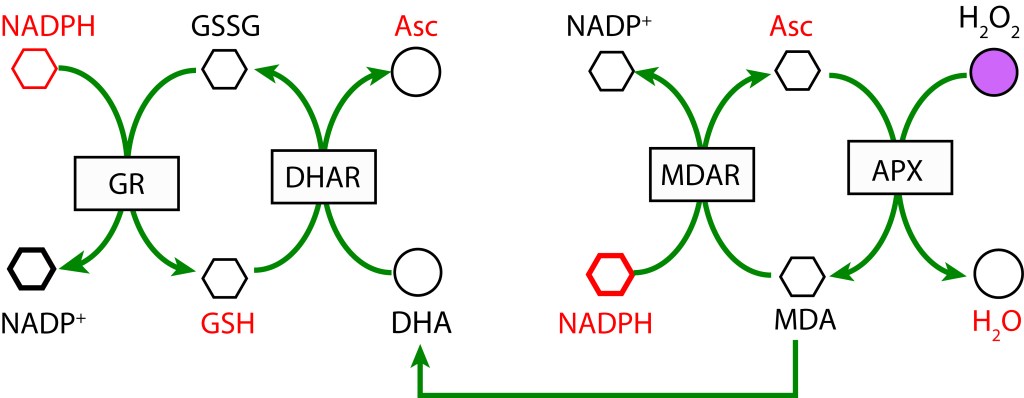
En un primer paso, MDA (Monodeshidroascorbato) se reduce a Ascorbato (ASC) usando NADPH como donante de electrones mediante la enzima MDAR (MDA Reductasa). En la etapa final MDA se regenera mediante la enzima Ascorbato Peroxidasa y transferencia de electrones al Peróxido de Hidrógeno para formar agua.
Aunque el ciclo se conoce como ciclo Agua-Agua, referido a que la fuente primaria de los electrones es agua (proveniente del ciclo de KOK), y que el producto final de la ruta también es agua, esta condición es común para las tres rutas correctivas.
RUTA ASCORBATO GLUTATION MEDIADA POR ASCORBATO PEROXIDASA
La ruta es activada tras la reducción de MDA (Monodehidroascorbato) a Ascorbato utilizando la enzima MDAR (Monodehidroascorbato reductasa) con NADPH como donador de electrones. MDA es regenerado mediante la acción de la enzima Ascorbato peroxidasa (APX) resultando en la reducción de H2O2 a H2O. Como radical, si no se favorece una rápida reducción, MDA dismuta a Ascorbato y Dehidroascorbato (DHA) .
En una segunda cadena de oxidoreducción, Dehidroascorbato se reduce a Ascorbato mediante la enzima Dehidroascorbato reductasa (DHAR), utilizando Glutatión (GDH) como agente reductor, el cual se oxida a su forma Glutatión disulfido (GSSG). Finalmente, GSH se regenera con NADPH como donor de electrones y la acción de la enzima Glutatión reductasa (GR).
ROL DEL ASCORBATO
ASC (forma salina dedl ácido Ascórbico, o Viamina C), es el principal antioxidante de cyanobacterias y plantas superiores. En adición a su función durante la fotosíntesis, participa como señalizador durante el proceso de la mitosis, forma parte del control de la función meristemal, contribuye al crecimiento radicular en plantas e interviene en la activación de la floración y durante la senecencia vegetal.
Cuatro rutas de síntesis de Ascorbato han sido descritas en plantas, con D-Glucosa, Myo-Inositol y pectinas de la pared celular como precursoras.
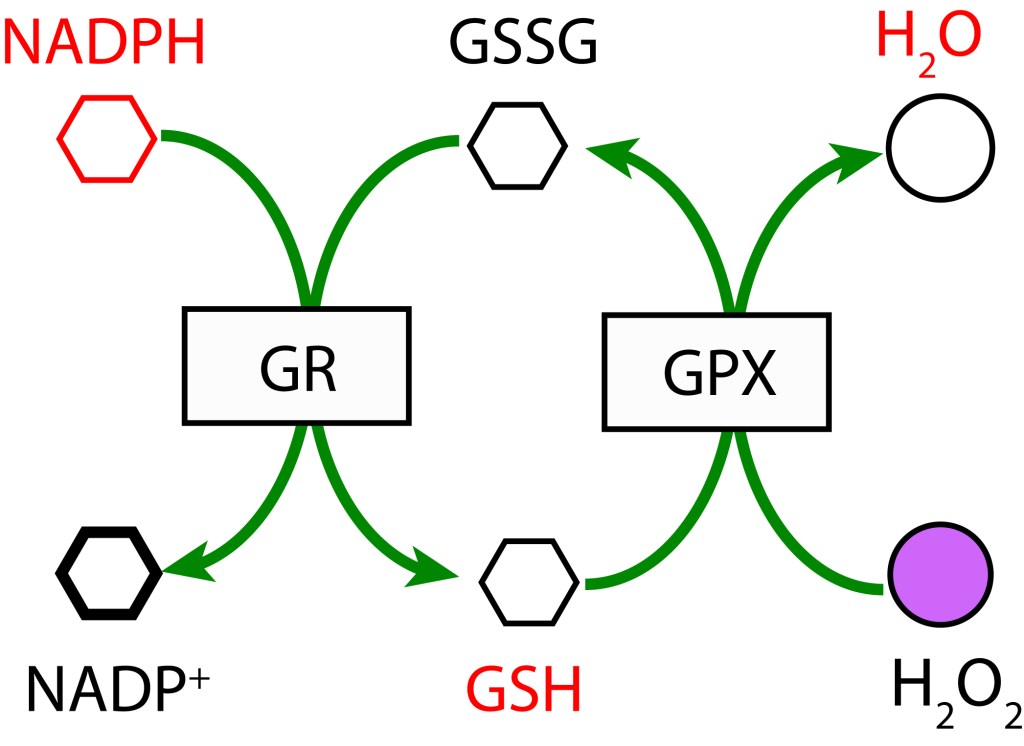
RUTA GLUTATION MEDIADA POR GLUTATION PEROXIDASA
En esta ruta, GSH participa como agente reductor actuando sobre H2O2 para producir H2O mediante la enzima Glutation Peroxidasa (GPX). GSH es regenerado a partir GSSG con NADPH como donor de electrones bajo la acción de la enzima Glutatión Reductasa (GR).
NPQ (NON PHOTOCHEMICAL QUENCHING)
Las rutas de protección expuestas previamente corresponden a protección electrónica de alta energía, presente en especies reactivas ROS.
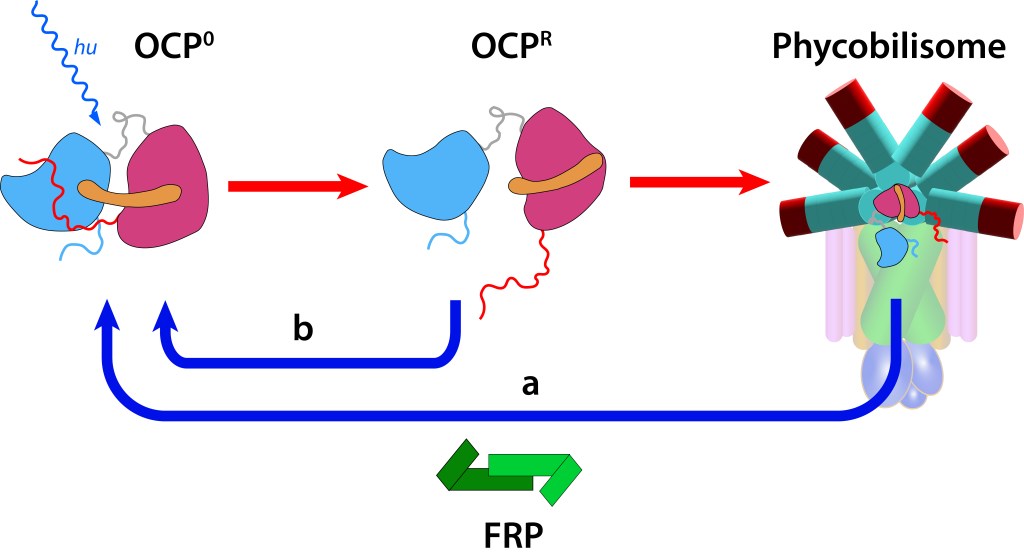
Existe adicionalmente una ruta de protección fotónica equivalente en función al ciclo de Xantophyllas propia de plantas superiores, destinada a disipar excesos de luz y calor en lo que se denomina Non-Photochemical Quenching” (NPQ), (Extinción No Fotoquímica), operando entre el nucleo del phycobilisoma y el centro de reacción RC impidiendo que fotones de alta energía degraden el fotosistema.
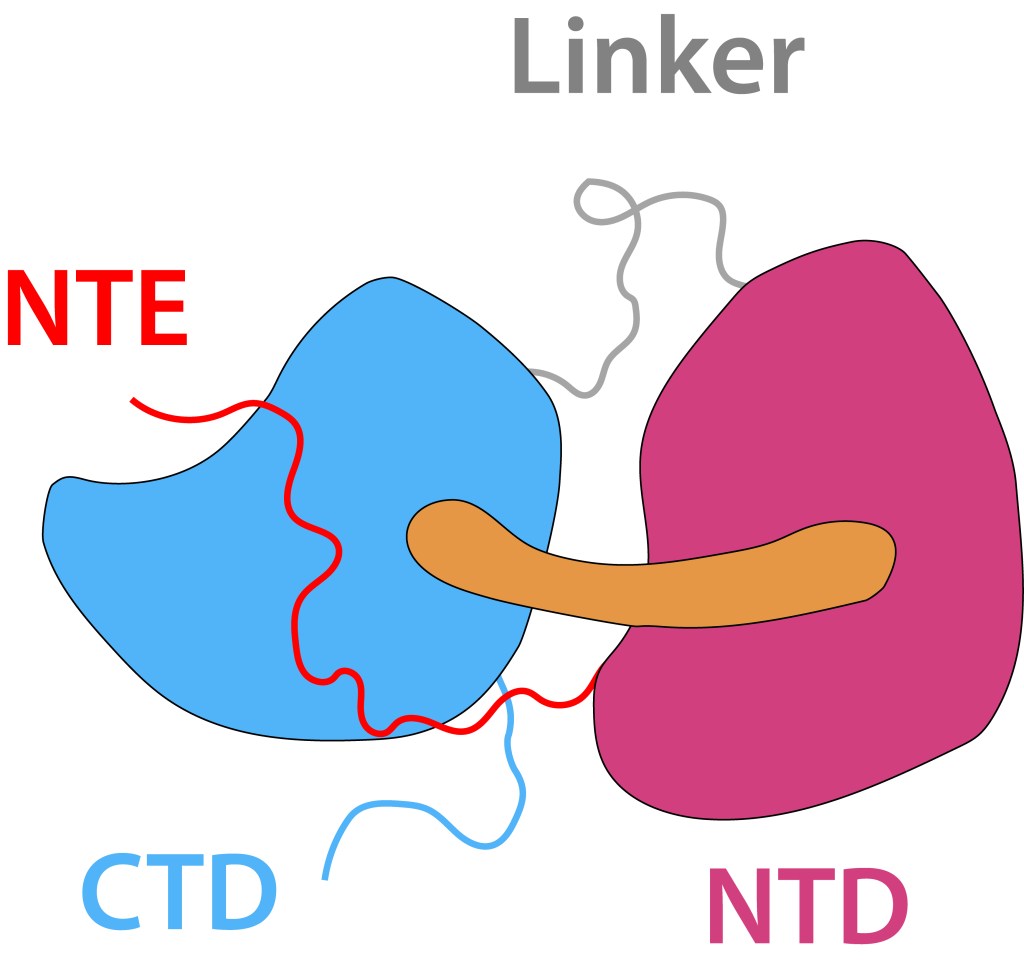
El core del sistema es la “Proteina Carotenoide Naranja”, “OCP”, ver Fig 4, molécula soluble en agua con un peso molecular de 35 kDa, dos dominios, C-Terminal (CTD) y N-terminal (NTD), separados por un linker que actúa a manera de bisagra entre los dominios, una terminal N extendida (NTE) contribuyendo a mantener CTD y NTD adyacentes en estado ground (OCPO) y un cofactor keto-carotenoide (Canthaxanthina).
Figura 4. Orange Carotene Proteine (OCP). Elementos principales
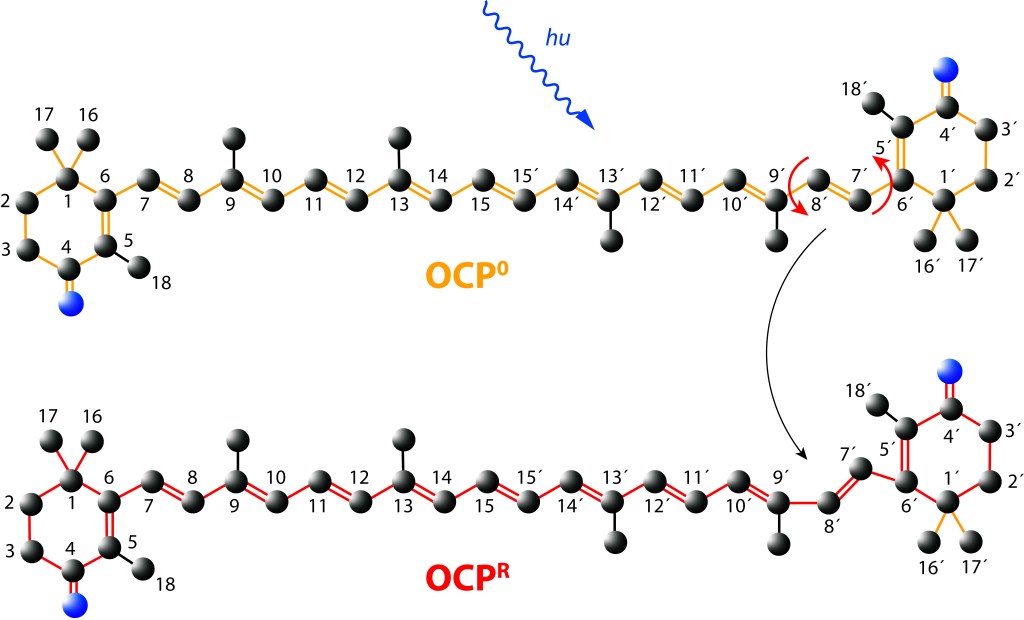
Ante la exposición a luz de alta energía, el cofactor Caanthaxanthina sufre cambios reversibles en su estructura molecular espacial, ver Fig. 5, con alteraciones en la absorción/emisión lumínica, pasando de naranja (ground) a roja en estado activado, loque define dos estados: OCP0 → OCPR.
Figura 5. Canthaxantica en estado Ground (OCP0) y estado exitado (OCPR)
Estos cambios generan a su vez alteraciones reversibles en OCP, ver Fig 5., incluyendo translocación del caroteno hacia el domino NTD y separación de los dominios N-terminal y C-terminal ver Fig.6.
Figura 6. Estructura y función OCP. (Modificado de Maksimov et.al, 2017).
En este estado OCPR actúa a nivel del nucleo del phycobilisoma impidiendo que alta energía se transmita hacia el centro de reacción, protegiendo de esta manera al fotosistema (fotoprotección NPQ).
Si la protección del fotosistema no es requerida, la proteina retorna a estado OCP0 mediante la acción del dímero FRP (Fluorescence Recovery Protein), lo que permite la reactivación de la función fotosintética.
BIBLIOGRAFÍA
Zolotereva, E., Polishuk,O. Chlororespiration as a Protective Stress-inducible Electron Transport Pathway in Chloroplasts. The Open Agriculture Journal • 14 Oct 2022
Slonimskiy, Y.B., Maksimov, E.G. & Sluchanko, N.N. Fluorescence recovery protein: a powerful yet underexplored regulator of photoprotection in cyanobacteria†. Photochem Photobiol Sci19, 763–775 (2020). https://doi.org/10.1039/d0pp00015a
Maksimov EG, Sluchanko NN, Slonimskiy YB, Mironov KS, Klementiev KE, Moldenhauer M, Friedrich T, Los DA, Paschenko VZ, Rubin AB. The Unique Protein-to-Protein Carotenoid Transfer Mechanism. Biophys J. 2017 Jul 25;113(2):402-414. doi: 10.1016/j.bpj.2017.06.002. PMID: 28746851; PMCID: PMC5529199.
Pagels, F.; Vasconcelos, V.; Guedes, A.C. Carotenoids from Cyanobacteria: Biotechnological Potential and Optimization Strategies. Biomolecules 2021, 11, 735. https:// doi.org/10.3390/biom11050735
Chukhutsina, V.U., Baxter, J.M., Fadini, A. et al. Light activation of Orange Carotenoid Protein reveals bicycle-pedal single-bond isomerization. Nat Commun13, 6420 (2022). https://doi.org/10.1038/s41467-022-34137-4
Las bacterias púrpura son un grupo de proteobacterias Gram negativas pertenecientes al órden Chromatiales, clase Gammaproteobacterias, presentes en cuerpos de agua anóxicos con alta iluminación.
A nivel bioquímico es un grupo muy versátil, pues dependiendo de las condiciones y recursos energéticos disponibles puede activar rutas de metabolismo fotoautótrofo, fotoheterótrofo, fermentativo, o respiratorio (anaerobio o aerobio).
Dada la importancia ecológica del grupo (particularmente como agente purificador de aguas residuales), su bioquímica ha sido ampliamente estudiada. La comprensión de su función fotosintética es de gran ayuda para abordar la dinámica de maquinarias más complejas, particularmente las correspondientes a cyanobacterias, algas y plantas superiores.
Antes de describir en detalle dicha maquinaria, vale la pena revisar algunos conceptos generales del fenómeno.
La fotosíntesis se define como el proceso de captura de energía lumínica y transpormación a energíia química, utilizando CO2 como fuente de carbono, estando presente en varios grupos bacterianos de bacterias, en algas y en plantas superiores.
El proceso comprende dos fases:
REACCIONES DEPENDIENTES DE LA LUZ: Ejecutadas de manera secuencial por diferentes complejos proteínicos transmembrana, parten de la captura de energía lumínica y se extienden hasta la síntesis de ATP y NADPH, ver Fig.1.
Fig.1. Etapas de las reacciones dependientes de la luz en la fotosíntesis.
REACCIONES NO DEPENDIENTES DE LUZ: Corresponde a un conjunto de reacciones de biosíntesis partiendo de agua y CO2, utilizando ATP y NADPH como fuentes e energía. Esta fase se realiza en el citoplasma mediante diversas rutas metabólicas reductoras, que dependiendo del grupo biológico pueden corresponder al ciclo de Calvin-Benson-Bassham (ciclo de las pentosas fosfato reductoras) , al ciclo reverso del ácido tricarboxílico o a la ruta del hidroxi-propionato, principalmente.
REACCIONES DEPENDIENTES DE LA LUZ
En las bacterias púrpura, las reacciones fotosintéticas dependientes de la luz ocurren en organelos especializados de la membrana interna celular, denominadas cromóforos o cromatóforos, ver Fig 2.
Fig 2. Cromatóforo en Rhodobacter sphaeroides. Fuente: Sener MK et al, 2007. Atomic level structure and function of an entire biological membrane: the photosynthetic unit of Rhodobacter sphaeroides. Proc Natl Acad Sci USA 104
Los cromatóforos surgen de invaginaciones de la membrana citoplasmática interna. Cada invaginación genera un volumen periplasmático confinado, limitado hacia el exterior por la membrana externa, ver Fig.3.
El reducido volumen de periplasma al interior del cromatóforo optimiza la generación de un gradiente de carga positiva, PMF, esencial para la síntesis de ATP.
Estrategias de confinamiento equivalentes se presentan en los Tilakoides de los cloroplastos, las invaginaciones tilakoidales de cyanobacterias, en el espacio periplasmático de heliobacterias e incluso en las crestas mitocondriales propias de células eukariotas.
Fig 3. Estructura de un cromatóforo
Las reacciones dependientes de la luz comprenden las siguientes etapas:
CAPTURA DE LUZ
EXITACIÓN DEL PAR ESPECIAL DE CLOROFILAS PRESENTES EN EL CENTRO DE REACCIÓN RC TIPO II (P870)
FLUJO DE ELECTRONES (Electron Transport Chain, ETC) a lo largo de una sucesión de reacciones de oxidoreducción a partir de la exitación y oxidación del par especial de clorofilas.
FUERZA PROTOMOTRIZ PMF (generación de un potencial protónico)
SÍNTESIS DE ATP (a partir de PMF como energía generatriz).
La Fig.4. resume a manera de diagrama de flujo el proceso fotosintético en este grupo, resaltando el colores los diferentes tiposde energía, incluyendo en rojo la energía lumínica (correspondiente a la captura y transporte de fotones), en verde la energía eléctrica, cuyo flujo se denomina ETC (Electron Transport Chain), en amarillo el flujo protónico (o flujo de iones H+) y finalmente en negro la depositación final de energía en la forma de ATP. Se omite en el diagrama la síntesis de NADPH, tema que se abordará más adelante, el cual requiere ATP y enzimas reductoras específicas. Con fines ilustrativos el centro de reacción RC y el complejo II se tratan como bloques independientes, si bién corresponden a un solo heterodimérico con sumbunidades L, M.
Fig 4. Diagrama de flujo reacciones fotosintéticas depenientes de la luz en bacterias púrpura
Como se aprecia en la figura, el complejo fotosintético de este grupo bacteriano se caracteriza por tener un solo fotosistema (Tipo II) , el cual es responsable de la reducción de quinonas y depositación de las mismas en el pool de quinonas, y un citocromo BC1 que activado por las quinonas reducidas responde por la generación de fuerza protomotríz (PMF) requerida para la síntesis de ATP y de la retroalimentación de electrones de vuelta al sistema. Finalmente, las reacciones dependientes de la luz, como también todas las maquinarias respiratorias, cierran el proceso mediante la enzima ATP Synthase, la cual es responsable de la síntesis de ATP, lo que permite en último término que la energía lumínica se transforme en energía química, se exprese en complejidad bioquímica y sostenga el proceso global celular operando adecuadamente .
Una vez acumulada la energía en forma de ATP entran en juego las reacciones no dependientes de la luz, anteriormente llamadas reacciones de obscuridad, que en este caso están representadas por el ciclo CBB. Este ciclo, activado por las moléculas mencionadas, permite capturar carbono a partir de CO2 y sintetizar glucosa. El ciclo se tratará de forma detallada en otra lectura. La fIG.5 exponede manera detallada la maquinaria fotosintética de bacterias púrpura . En la esquina superior izquierda de la figura se ilustra la invaginacion de la membrana interna y el espacio periplasmático donde tiene lugar el proceso.
Fig 5. Bacterias púrpura: Aparato fotosintético
REACCIONES DEPENDIENTES DE LA LUZ
CAPTURA DE LUZ (LIGHT HARVESTING)
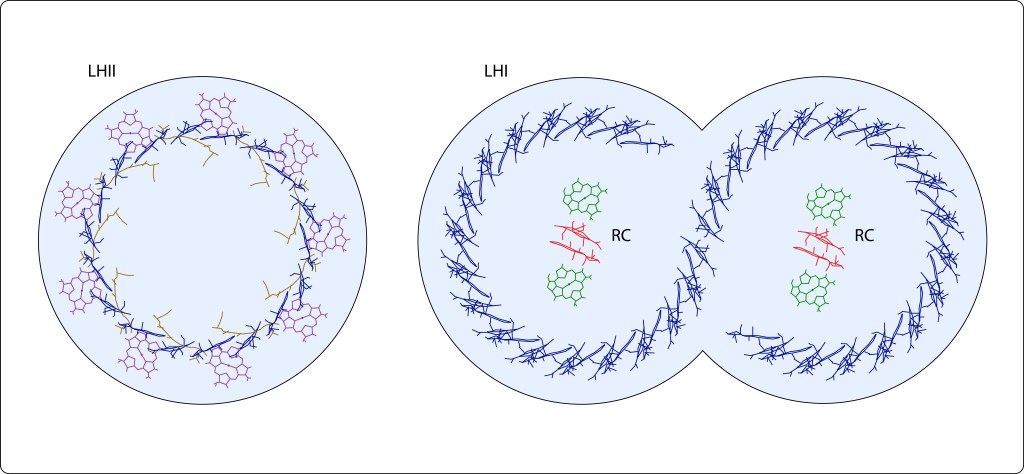
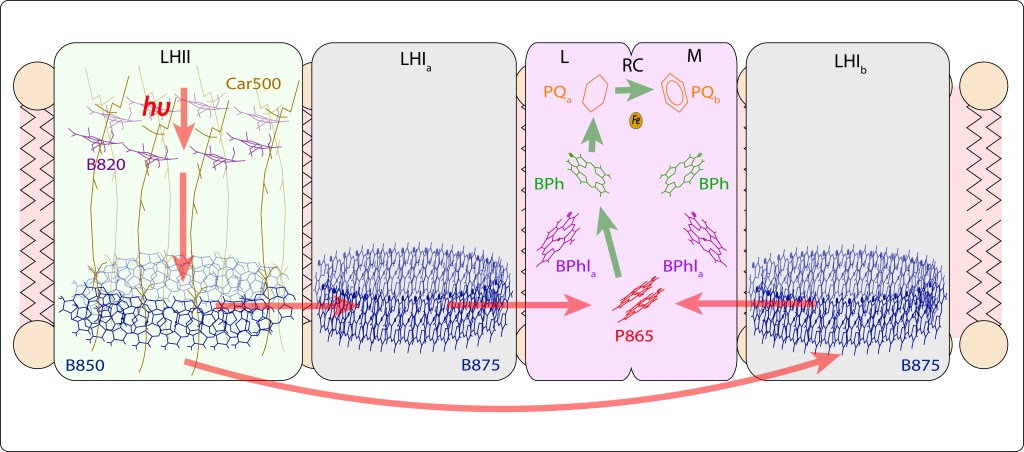
En bacterias púrpura, la energía lumínica exita pigmentos fotosintéticos, carotenos y clorofilas, geométricamente distribuídos al interior de dos tipos de complejos proteicos transmembrana denominados complejos antena LHII y LHI, ver Fig 6.
Fig 6. Vista superior de los complejos antena de Bacterias Púrpura. LHII (izquierda) contiene 9 carotenos (café) en empalizada , 9 Bchla´s paralelas y 18 Bchl´s perpendiculares a la membrana plasmática. LHI dimérico en forma de “S” contiene 36X2 clorofilas dispuestas perpendicularmente a la membrana plasmática. Círculos en azul claro representan las proteinas transmembrana correspondientes a cada monomero.
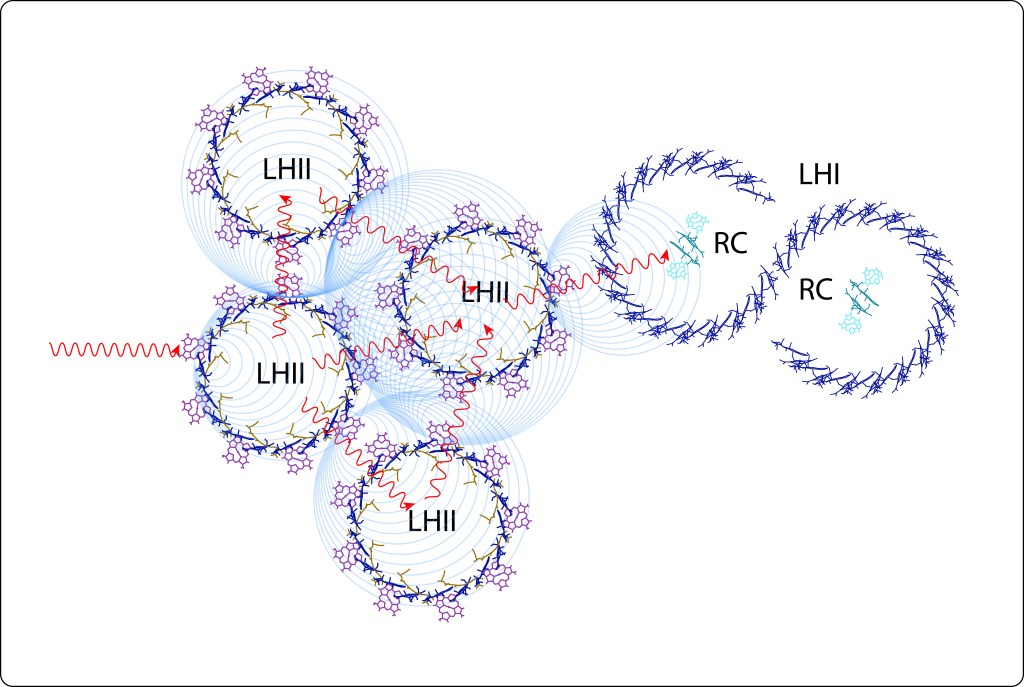
Mediante resonancia no radiativa (Resonancia de Förster) y coherencia cuántica, la energía capturada en LHII es transferida a LHI, para finalmente ser transladada a un par especial de bacterioclorofilas, denominadas Special Pair, localizadas en el Centro de Reacción, RC, ver Fig 7.
Fig 7. Flujo de energía desde los Complejos Antena LH hasta en centro de reacción RC.
La transferencia radiativa implica una disminución gradual de la energía en el tránsito desde LHII hasta LHI.
2. EXITACIÓN DEL PAR ESPECIAL DE CLOROFILAS
El par especial P870 en RC exitado por un valor de energía superior al valor de exitación de su donor en LHI (B875)., ver Fig. 8, lo cual se debe probablemente al efecto combinado de coherencia y superradiancia cuánticas.
Fig 8. Energías y rutas de exitación en los Complejos Antena LH hasta en centro de reacción RC. Transcrito de Proc. Natl. Acad. Sci. USA 95 (1998)
3. FLUJO DE ELECTRONES ETC
El flujo de energía lumínica, indicado con flechas en rojo en la Fig 8., tiene lugar vía LHII → LHI → P870 en RC, mientras que la cadena de transporte de electrones ETC, flechas en verde, ocurre desde el par especial hasta PQ b en el centro de reacción RC. Ver Fig.9.
Fig 9. Flujo de energía en los complejos antena de Bacterias Púrpura. Se incluye la transferencia de LHI a RC, y la cadena de transporte en RC, tema amplado más adelante. Las subunidades LHIa y LHIb en la imagen corresponden a un único complejo LHI en “S” , ver también Fig 3., rodeando el centro de reacción RC . Note que el complejo proteico de RC está formado por un dímeero con subunidades L, M.
NIVELES DE ENERGÍA
Los pigmentos fortosintéticos presentan, dependiendo de la energía incidente, dos niveles de exitación: 1° y 2° estados en singlete, ver Fig 10.
Fig 10. Niveles de energía en la molécula de clorofila. Dependiendo de la luz incidente, el estado de exitación puede ser 2° singlete (banda azul), o 1° singlete (banda roja). Fuente : Biochemistry & Molecular Biology of Plants.2015, John Willey & Sons.
Electrones en 2° singlete se relajan rápidamente de manera expontánea a 1° singlete por conversión interna (IC) y liberación de calor, mientras que los de 1° singlete (en el caso del par especial de clorofilas) pueden tener dos destinos:
Retorno a GND por fluorescencia
Liberación desde el par especial vía REDOX (reducción de BPha como primer aceptor de electrones) , iniciandose con este evento la cadena de transporte de electrones ETC. La ruta tomada dependerá de los requerimientos fisiológicos de ATP. Alta demanda favorecerá ETC, caso contrario la molécula retornará a GND.
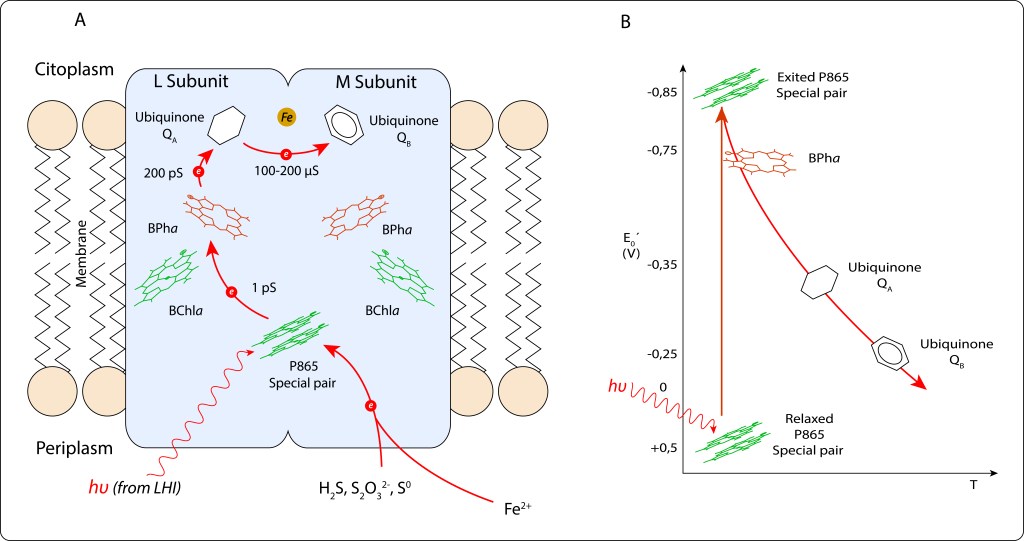
Ante demanda de ATP, el electrón cedido por el par especial inicia una serie de reacciones REDOX en cadena, ver Fig 10.
P865 → BPha → QA → QB.
Cofatores adicionales, BChla y Fe2+ , indicados en la figura 11., no participan directamente en la cadena de transporte, su función es estabilizar la carga local para facilitar la unidireccionalidad de la cadena de reacciones.
Fig 11. Cadena de transporte de electrones en RC. A. Pigmentos y moléculas involucradas en el transporte. B. Niveles de energía correspondientes a cada etapa.
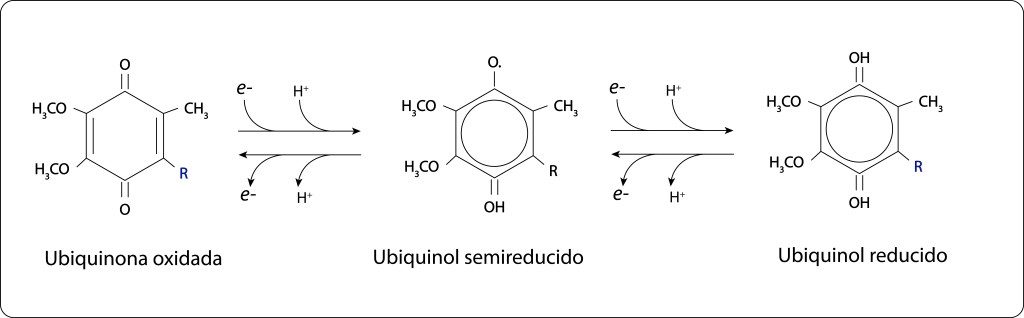
El aceptor final en RC, Q b , requiere dos electrones para reducirse completamente, ver Fig 12., de modo que el par especial debe ceder secuencialmente dos electrones , lo que implica captura de dos fotones por parte de los complejos antena y consecuentemente utilización de los dos electrones que proceden del citocromo C en una ruta cíclica.
Fig 12. Reducción de Ubiquinona a Ubiquinol, pasando por Ubiquinol semireducido.
La plastoquinona completamente reducida migra al pool de quinonas y mediante la adquisición de 2 cationes H+ procedentes del citoplasma se convierte rapidamente en Plastoquinol . El confinamiento de plastoquinonas y plastoquinol al interior de la membrana citoplasmática interna es posible gracias a su alta hidrofobicidad molecular, de manera que los fosfatos de la bicapa fosfolipídica actuan como barrera de retención manteniendo las quinonas y quinoles en el pool a disposición del citocromo Bc1.
4. FUERZA PROTOMOTRIZ PMF
Ubiquinol se convierte entonces en el portador de energía a ser utilizado por el citocromo bc1 para la generación del gradiente protónico requerido para la síntesis posterior de ATP.
El complejo citocromo bc1 , al igual que b6f en cianobaterias y en mitocondrias de células eucariotas, se caracteriza por contener moléculas Heme como cofactores responsables del transporte de electrones.
En bc1, tres Heme participan en el transporte de electrones, dos en Cyt b y una en Cyt c1; lo que contrasta con el centro de reacción RC, donde los cofactores responsables del transporte corresponden a clorofilas, phaeophytinas y quinonas.
En Heme, un átomo de Hierro Fe2+ es quelatado de manera similar a la quelatación de magnesio Mg2+ en las clorofilas y bacterioclorofilas, ver Fig 13.
Fig 13. Esquema comparativo de las moléculas de Bacterioclorofila a y Heme.
Mientras que en RC los electrones cedidos por el par especial de clorofilas corresponden a electrones π de la cadena resonante de enlaces conjugados, en heme los electrones provienen del ión ferroso Fe2+ al pasar a estado férrico Fe3+.
Heme y clorofila comparten una misma ruta de síntesis metabólica, partiendo del ácido δ-aminolevulínico (ALA) hasta el subproducto protoporfirina IX. De ahí en adelante los agentes quelatantes y demás diferenciadores definen el producto final.
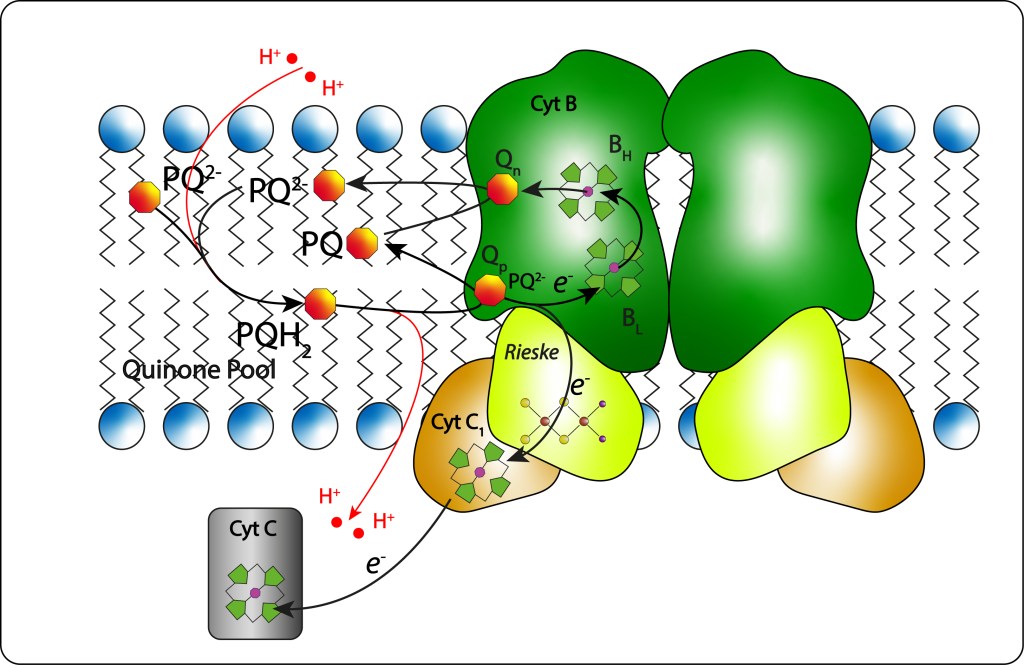
El Ubiquinol (QH2) exportado desde el centro de reacción al pool de quinonas es redireccionado al sitio Qp del CytB, ver Fig 14, donde es totalmente oxidado.
Fig 14. Cytochrome BC1. Función y diagrama general
Un primer electrón es capturado por el cofactor Fe2S2 del complejo Rieske, ver Fig 15., de allí pasa al citocromo C1 y luego al Cyt C2 soluble que lo lleva de nuevo al centro de reacción RC para reducir el par especial de clorofilas, pudiendo reiniciarse así un nuevo ciclo de transporte de electrones.
Fig 15. Complejo Rieske. Uno de los dos hierros es coordinado por dos residuos His definiendo su alto potencial Redox.
El segundo electron sigue una ruta diferente: Heme Bl (bajo potencial) -> Heme BH (alto potencial) → quinona Q (localizada en un segundo sitio de CitB: Qn) reduciendola parcialmente (Q → Q–) , de manera que la reducción completa de Q implica la reducción de dos ubiquinoles.
La figura 16 ilustra los potenciales Redox en bacterias púrpura y bacterias verdes del azufre (para fines comparativos) a lo largo de la cadena de transporte de electrones, incluyendo las etapas que tienen lugar en RC y las etapas en Cyt bc1. Note que en bacterias púrpura la reducción de NAD+ de manera directa por la vía de fotosíntesis, por su alto valor de potencial.
Como se puede observar, de manera global los potenciales Redox de bacterias púrpura son menores comparados con los potenciales de bacterias verdes del azufre.
Esto define para el sistema de bacterias púrpura un poder oxidante fuerte, reductor débil, mientras que GSB se define un poder oidante débil, reductor fuerte.
la importancia de esta diferencia en poder oxidoreductor entre uno y otro grupo biológico se entenderá más adelante, al abordar las cyanobacterias, un grupo biológico que por endosimbiosis bioquímica (término personal propuesto en estas lecturas) integró los dos fotosistemas en una sola maquinaria fuertemente oxidante, fuertemente reductora, capaz de oxidar H2O y capaz de reducir NAD+ de manera autónoma, lo que en su momento evolutivo defino un nuevo curso para la vida en el planeta.
Figura 16. Potenciales redox en bacterias purpura y verdes del azufre
5. SÍNTESIS DE ATP
Como resultado de la oxidación del Ubiquinol, dos hidrogeniones H+ son exportados al periplasma contribuyendo a generar el gradiente PMF requerido para la síntesis de ATP.
El citocromo BC1 actua entonces como una bomba de protones, desde el citoplasma al periplasma, y el PMF generado actúa como habilitante de la ATP Synthase, último complejo de la cadena de reacciónes dependientes de la luz.
El diferencial de iones H+ entre periplasma y citoplasma, PFM, es utilizado por ATP synthase para la síntesis de ATP.
Si el Citocromo BC1 es una bomba de protones, ATP synthase es una verdadera ensambladora rotativa de tres tiempos, admisión, compresión y expulsión, con un rotor dotado de tres sitios activos separados 120° entre sí. La rotación es habilitada gracias al flujo de protones procendentes de PMF a travez de la ATP Synthase.
El primer sitio activo, o sitio de admisión, recibe ADP y radicales fosfato. El sitio de compresión inserta mediante ataque nucleofílico el fosfato en la molécula de ADP para generar ATP, y finalmente en el sitio de expulsión el ATP se libera al citoplasma quedando a disposición de los procesos bioquímicos celulares, y en particular a las cadenas fotosintéticas no dependientes de la luz, que en el caso de las bacterias púrpura, corresponden al ciclo de Calvin-Benson.
Esta enzima no es exclusiva de las bacterias púrpura, y está presente prácticamente en todos los organismos vivientes, desde arqueas hasta plantas superiores y animales, y siempre su función es la síntesis de ATP, utilizando el gradiente electroquímico de protones H+ , aunque en algunos casos también utiliza el gradiente de iones Na+ (Fuerza Sodio-motriz) , y raramente K+.
6. SÍNTESIS DE NADPH – NADH
Como se ilustró en la Fig.16., el centro de reacción P870 característico de bacterias púrpura NO es lo suficientemente reductor para reducir directamente NAD⁺, de modo que se debe contar con una maquinaria adicional para producirla.
Existen varias rutas reductoras principales:
NADPH vía electrones provenientes de Cyt bc1:
Por esta ruta, parte de los electrones provenientes de Cyt bc1 via complejo de Rieske no retroalimentan RC sino que son redireccionados a Fdox para su reducción:
Fdox +e– → Fdred
A continuación, la enzima FNR (ferrodoxina-NADP+ reductasa), cataliza la reacción
Fdored + NADP+ + H+ → Fdox + NADPH
NADH víaElectrones provenientes de donores orgánicos (Succinato, lactato):
En esta ruta, dado el bajo poder reductor de la maquinaria fotosintética y alto potencial Redox del par NAD+/NADH, se habilita el complejo II (Complejo SDH) de la máquinaria respiratoria (Succinato deshidrogenasa), permitiendo que succinato (o lactato) reduzcan quinonas a ser redireccionadas no hacia Cyt bc1 sino hacia el complejo I de la cadena respiratoria. Utilizando ATP-asa (ATP-Sinthase inversa) se incrementa el gradiente protónico en el espacio periplasmático lo que genera un flujo protónico inverso que a su vez permite que los electrones fluyan “Up Stream” en la cadena de ferrodoxinas hasta finalmente reducir NAD+. Ver Fig.17. Este proceso ocurre en obscuridad, de manera que el aparato fotosintético (módulos en gris en la figura) permanecen inactivos. Con propositos comparativos se anexa como Fig.18 el complejo fotosintético previamente presentado en la Fig 4. Vale la pena resaltar la función de “Switch” o conmutador de energia de l complejo II, el cual no solo alimenta alternativamente el Complejo I en la síntesis de NADH o el pool de quinonas en el proceso fotosintético sino que también invierte el flujo protónico, convirtiendo ATP-Synthase en ATP-Asa, forzando a un flujo protónico de citoplasma a pariplasma, y tambien invirtiendo este flujo en el complejo I, lo que finalmente lleva a sintetizar (y no consumir) NADH.
Figura 17. Síntesis de NADH en bacterias púrpura fotosintéticas via complejos de la cadena respiratoria.
Figura 18. Diagrama de flujo reacciones fotosintéticas dependientes de la luz en bacterias púrpura
NADPH vía transhidrogenasas.
Las transhidrogenasas son enzimas que transfieren hidruros (H⁻) entre NADH y NADP⁺ para producir NADPH, siendo en la práctica un intercambio de poder reductor entre estos dos cofactores:
NADH + NADP+ ↔ NAD+ + NADPH
Esta ruta es particularmente útil cuando:
Hay exceso de NADH (por respiración o metabolismo de sustratos orgánicos).
Hay demanda de NADPH para biosíntesis (fijación de CO₂ cia CBB, síntesis de lípidos, etc).
REACCIONES NO DEPENDIENTES DE LA LUZ
En bacterias púrpura, al igual que en cyanobacterias y plantas superiores, el ciclo de Calvin-Benson es el receptor de facto del ATP producido en la fotosíntesis. Este ciclo forma parte integral de la maquinaria fotosintética, y corresponde a lo que se denomina reacciones no dependientes de la luz.
El ciclo de Calvin-Benson merece una lectura aparte, luego no se detalla en el presente blog.
Para abordar el tema de la fotosíntesis, un buen punto de partida puede ser su historia evolutiva.
La fotosíntesis, entendida como la combinación del aprovechamiento directo de la energía lumínica y la incorporación de carbono en la maquinaria viviente a partir de CO2, solo se presenta en bacterias, algas y plantas superiores. En arqueas, la otra rama evolutiva de la vida, existe un mecanismo de captura directa de luz pero nunca incorporando carbono a partir de CO2.
En cuanto a las bacterias, la maquinaria fotosintética debió evolucionar a partir de transferencia horizontal desde maquinarias fosforilativas propias de bacterias heterótrofas.
No hay concenso en la comunicad científica en cuanto al origen evolutivo de la fotosíntesis; quizas los primeros pigmentos fotosintéticos fueron incorporados inicialmente como un mecanismo de protección UV (terrabacterias ancestrales colonizadoras de tierra firme con alta exposición al UV) , en un mecanismo similar a como son utilizadas las Xantofilas actuales, para luego ser aprovechados como elementos resonantes de la energía lumínica en vías similares a las actualmente utilizadas en la fotosíntesis.
Dos tipos de complejos fotosintéticos están presentes en los organismos fotosintétizadores: PSII, complejo áltamente oxidante, capaz de oxidar el agua, y PSI, muy buen reductor, capaz de reducir moléculas acumuladoras de energía.
Las maquinarias fotosintéticas en los diversos grupos evolutivos incluyen uno o los dos fotosistemas, PSII o PSI, encargados de la captura primaria de energía, y un complejo citocromo, BC1, análogo a B6f de los aparatos de fosforilación oxidativa, gestor de la fuerza protomotríz que luego será aprovechada por la ATP sintasa para generación de ATP.
Un salto evolutivo enorme lo dieron las cyanobacterias, quienes, seguramente por transferencia horizontal, se apropiaron y pusieron a operar de manera armónica los dos fotosistemas, con el citocromo compartido a manera de pivote del complejo fotosintético.
Complementaridad !!!!
Un elemento decisivo en la evolución de la vida.
Endosimbiosis a nivel bioquímico, lo que dió como resultado una sinergia de función entre PSI y PSII, siendo la evolución convergente mas impactante en la historia de la vida.
Surgen varios elementos de reflexión sobre el papel de la evolución divergente (Darwinismo) y la evolución convergente (Margulismo), y me atrevo a resumirlo en una frase:
“Darwin propone, Margulis dispone”.
Se requiere una oferta amplia de opciones evolutivas, proporcionada por las dogmáticas mutaciones al azar de Darwin, para que Margulis, independientemente de la indiscutible selección natural, tome opciones y las combine de manera complementaria en nuevos y más eficientes fenómenos simbióticos de alto impacto.
Darvin y Margulis permitieron entender la evolución ya no como ramas darwinistas que se desprenden de un único tronco común para competir y nunca más colaborar, tampoco raíces que de manera colabortiva convergen a dicho tronco, mucho menos raices tronco y ramas formando un “bonito” árbol de la vida, más bien una intrincada red de ramas y troncos que se separan, se unen en opciones cada más complejas, compiten, colaboran, luchan, sobreviven, fallan, se equivocan, desaparecen, comparten información, optimizan procesos, etc, formando una “red evolutiva” a manera de red neuronal gestora de la biodiversidad planetaria.
Así pues, los saltos evolutivos de mayor impacto se dan por mecanismos simbióticos, pero siempre requiriendo de una amplia oferta de opciones de evolución divergente.
Ensamblar una máquina altamente oxitadiva con una máquina fuertemente reductora es proporcionarle a la vida un motor turbocargado, capaz de oxidar el agua, un inagotable recurso energético, y así extraerle sus electrones, transportadores de la energía, y los protones, que luego aportarán a la fuerza protomotriz requerida por la ATP Sintasa , convirtiendo a sus portadores (cyanobacterias) en el gran protagonista de la trama de la vida hace 2,500 millones de años.
Y es que las cyanobacterias se convirtieron en EL JUGADOR (en mayúsculas), con el poder de “envenenar” la atmósfera con oxígeno, el inevitable desecho del agua, llevándose en su momento por delante el 95% de la vida en el planeta en un fenómeno conocido como Gran Evento de Oxidación (Huronian GOE), adquirieron el poder de actuar sobre la geología planetaria a gran escala, permitiendo que el hierro disuelto se oxidara de su forma ferrosa soluble a su forma férrica insoluble para generar las famosas bandas geológicas de hierro; poder para cambiarle el color a la tierra vista desde el espacio, de un rojizo propio de una atmósfera de metano y Co2 a un color azul propio del oxígeno, pasando en el medio por un color blanco al despojar al planeta de su abrigo de invierno (atmósfera de CO2 y Metano) convirtiendolo en una bola de nieve (Snow Ball), pero también aquiriendo el poder de ser punto de inflexión en la evolución de la vida.
Varios grupos de bacterias actuales operan con el fotosistema PSII, varios con PSI, pero fueron las cyanobacterias quienes aprovecharon el poder dual, generando una amenaza , …. y una oportunidad, para la vida en el planeta.
En respuesta a la amenaza, algunas arqueas Asgard y un linaje particular de proteobacterias aeróbicas, quizas aeróbicas facultativas, resolvieron la amenaza del oxígeno atmosférico en en un nuevo evento de simbiosis, primero con tímidos coqueteos sintróficos entre pares complementarios para llegar finalmente en una íntima unión que hoy conocemos como Eukariogénesis, células con núcleo.
A3 (Entagling, Engulfing Endosymbiosis) resume el fenómeno, que finalmente convierte al par simbiótico en organelos especializados , mitocontrias de orígen bacteriano, nucleos arqueanos.
De manera que cada célula de nuestro organismo viene a ser habitación de una arquea y una bacteria !!! . Somos en esencia bacterias y arqueas conviviendo en armonía !!!!
Y luego vino mas adelante un tercer evento simbiótico (otra vez Margulis, otra vez complementaridad), lo que permitiría incorporar una cyanobacteria (que luego se especializaria en forma de cloroplastos) al interior de la maquinaria eukariota, dando lugar a las algas, y mas adelante a las plantas superiores.