

Figura 1. Arreglo de pigmentos de una subunidad tubular en el cuerpo del Clorosoma de Bacterias verde sulfurosas
El clorosoma es el organelo responsable de la captura de energía lumínica en las maquinarias fotosintéticas de tres grupos de bacterias Gram (-) :
- Bacterias Verdes Sulfurosas (GSB), representadas por el género Chlorobium ( C. tepidum), y los géneros Chlorobaculum, Prosthecocloris y Cloroherpeton.
- Bacterias Fotótrofas Anoxigénicas Filamentosas (FAP), anteriormente bacterias verdes no sulfurosas, representadas por el género Chloroflexus (C. aurantiacus), y los géneros Roseiflexus, Oscilochloris, Heliothrix y Chloronema.
- Acidobacterias, con un único género fotosintético descrito a la fecha, Chloroacidobacterium.
Equivalente en función a los complejos LHII y LHI de bacterias púrpura y a los Ficobilisomas de las Cyanobacterias, los clorosomas son los orgenelos responsables de capturar la energía lumínica y redireccionarla al centro de reacción RC donde la separación de cargas tiene lugar.
Los Clorosomas se diferencian de los demás complejos antena por su gran tamaño (formacilindrica de corte elipsoidal con una longitud entre 100 y 200 nm, 50 nm de anchura, 25 nm de altura), por la ausencia de una matriz proteica que de soporte y organización a los pigmentos fotosintéticos y por el elevado número de pigmentos por unidad (250,000 moléculas de bacterioclorofilas c, d y e por clorosoma).
En ausencia de la matriz proteica, los pigmentos se autoensamblan en agregados paracristalinos, ver Fig.1., lo que los hace objeto actual de investigación en nanotecnología (fotosíntesis artificial y electrónica molecular).
El compacto agregado de pigmentos fotosintéticos proporciona un acoplamiento exitónico intramolecular muy fuerte. Este elemento, sumado a la enorme cantidad de moléculas involucradas, son la base de la alta eficiencia en captura de energía lumínica, al punto que unos pocos fotones incidentes por pigmento por día le permiten a las bacterias prosperar en ambientes anóxicos de extremada baja luminosidad (hasta 100 metros de profundidad en el Mar Negro), siendo considerados a la fecha los más eficientes complejos antena encontrados en la naturaleza.
ANATOMÍA DEL CLOROSOMA
El clorosoma está conformado por un cuerpo, responsable directo de la captura de energía, una capa conectora base (Baseplate), y en el caso de GSB, una capa proteica especial, FMO, (Fenna-Matthews-Olson), responsable de transmitir la energía al centro de reacción RC, ver Fig.2.

Figura 2. Estructura general de un clorosoma
Los dos principales pigmentos presentes en el clorosoma corresponden a BChl c y BChl a , en cuerpo y baseplate respectivamente, ambos con un precursor común, Chlorophyllide a, pero con diferencias importantes en su estructura molecular como se ilustra en la fig 3 y en la tabla 1.
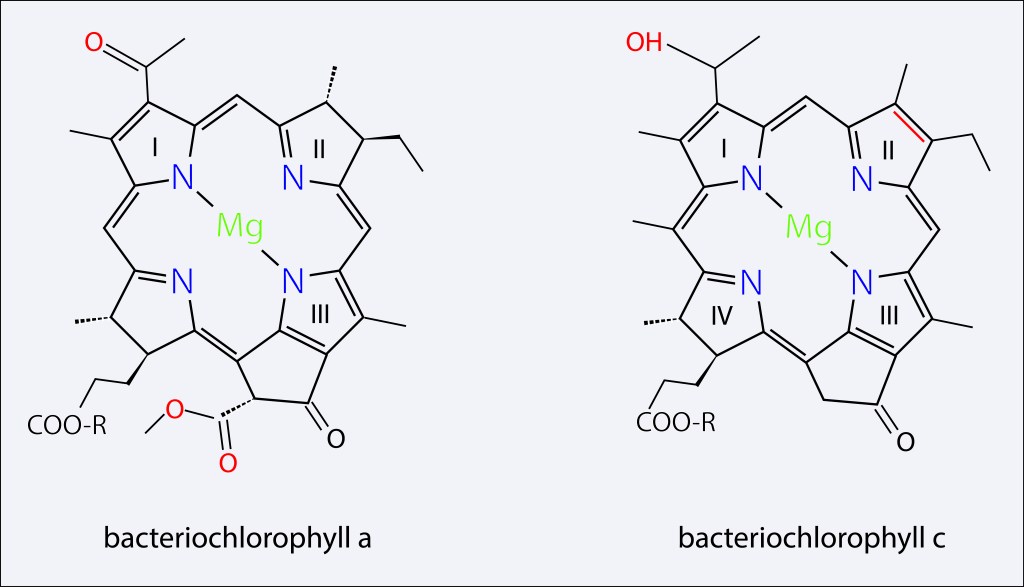
Figura 3. Pigmentos presentes en el clorosoma
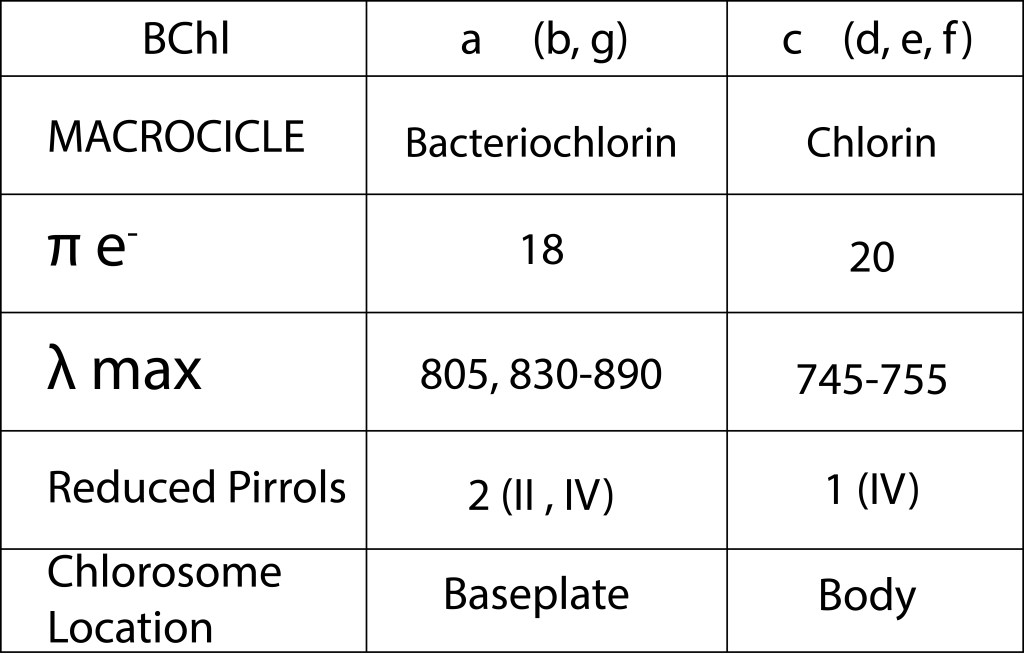
Tabla 1. Características de los pigmentos mas representativos presentes en el clorosoma
Los pigmentos fotosintéticos responsables de la captura primaria de energía corresponden principalmente a bacterioclorofilas del tipo CBhl c, en un número entre 60,000 y 250,000 unidades por clorosoma, cantidad que depende de la especie y de la disponibilidad de energía lumínica del entorno. Chlorobium tepidum contiene entre 200 y 250 clorosomas, para un total de cerca de 50 millones de moléculas de BChl c, lo que equivale a cerca del 30% del carbono celular.
Al interior del cuerpo los pigmentos se autoensamblan en subunidades tubulares, ver Fig1., mediante enlaces de coordinación C-31OH-Mg, ver Fig 4; absorben energía a 750 nm y por resonancia de Förster y coherencia cuántica direccionan la misma a la Base plate a 795 nm .
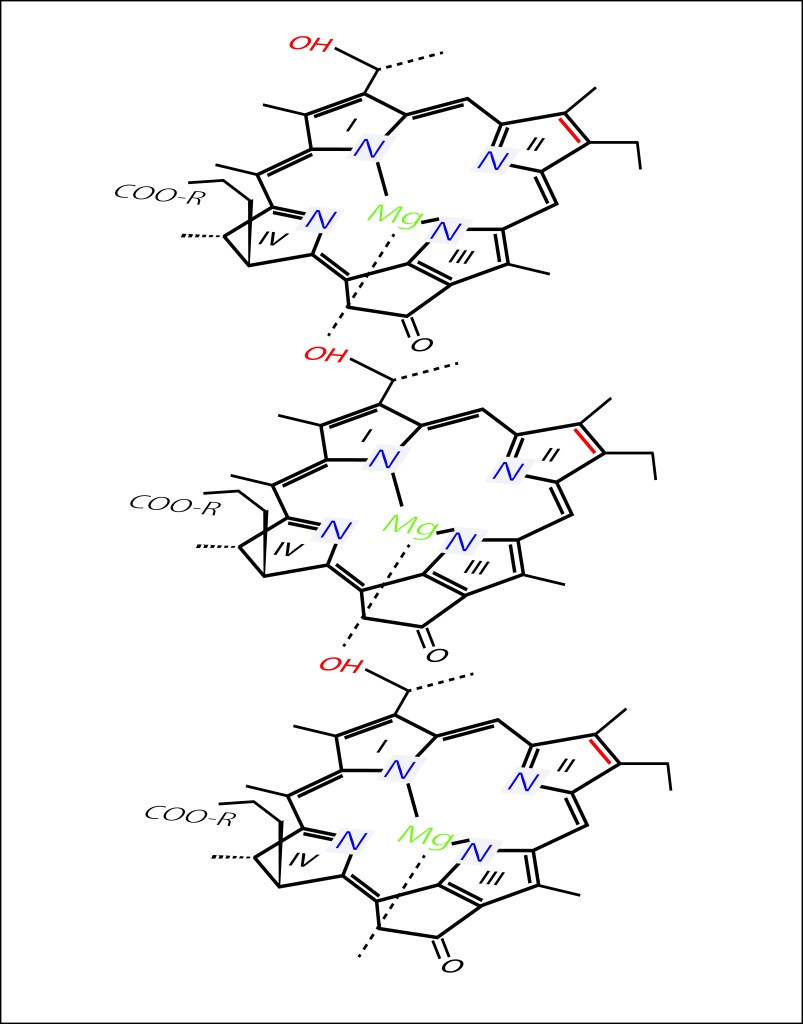
Figura 3. Apilamiento de moléculas BChl C bajo una estructura “Parallel Stack”. Otros arreglos son posibles (Piggy-back dimer model, Anti-parallel monomer stack model y Syn-anti monomer stack model). Visitar: Photosynthesis Research · June 2013. DOI: 10.1007/s11120-013-9869-3 · Source: PubMed. Para todos los casos, un enlace de coordinación C-31OH-Mg es el elemento enlazante de la cadena. Para los demás arreglos,
El conjunto de tubulos está rodeado por una membrana monofosfolipídica de geometría cilíndrica y corte elipsoidal poblada de proteinas clorosómicas transmembrana (CSM) pertenecientes a las familias CsmA, CsmB/CsmF, CsmC/CsmD y CsmH/I,J,X. La función de estas proteínas es contribuir a la estabilización y organización estructural de los pigmentos, y en el caso de CSMJ protejer el sistema mediante disipación de energía lumínica (quenching ).
En adición a las bacterioclorofilas, el cuerpo contiene carotenos que participan en la captura y transmisión de energía, y quinonas , particularmente clorobiumquinona, que operan como agentes protectores quencher.
En cuanto a la capa basal del clorosoma, o Base Plate, está conformada por proteínas CSMA ligadas a la capa monofosfolipídica en una estructura paracristalina 2D. las proteínas a su vez contienen pigmentos BChl a y probablemente carotenos. Cada monómero de CSMA está compuesto por dos hélices alpha, donde la hélice N-Terminal (residuos 6-36) se encuentra inmersa en los lípidos mientras la hélice C-Terminal (residuos 41-49) se asocia a la proteina FMO.
Hélices Alpha de monómeros adjuntos establecen interacciones hidrofóbicas que favorecen conformaciones diméricas, las cuales son la base del arreglo molecular de la Base Plate.
Finalmente, la capa FMO, que conecta la Baseplate con en centro de reacción en bacterias verde sulfurosas, es un trímero proteico con simetría de orden 3, compuesto por monómeros idénticos, ver Fig 4.

Figura 4. Disposición de los monómeros CSMA en FMO de Chlorobaculum tepidum. Fuente: https://www.ks.uiuc.edu/Research/fmo/
Cada monomero de CSMA contiene en su interior siete bacterioclorofilas BChl a, ver Fig 5, aunque en Chlorobaculum tepidum y otras bacterias verdes sulfurosas se ha encontrado evidencia de una octava BChl a más débilmente ligada, que parece cumplir un papel de acoplamiento con la Baseplate. Estos pigmentos son los responsables finales del tránsito de energía desde el complejo antena hasta el centro de reacción.

Figura 5. Disposición de las BChl a al interior de los monómeros CSMA. Fuente: Energy-scales convergence for optimal and robust quantum transport in photosynthetic complexes. The Journal of Chemical Physics · April 2011. DOI: 10.1063/1.4856795
Para concluir, es de anotar que FMO fué el primer complejo proteina-pigmentos en ser estudiado y entendido. Gran parte de lo que se conoce hoy en día de la interacción entre pigmentos y proteinas proviene de los estudios de dicho complejo. FMO fué igualmente el primer caso en que se demostró la ocurrencia de coherencia cuántica como fenómeno fundamental en la transmisión de energía hacia el centro de reacción. FMO solo está presente en especies con centros de reacción tipo I, carcterizados por la presencia de ferredoxina como aceptor de electrones en la cadena de transporte de electrones ETC, tema que se tratará en las próximas lecturas.
