

Fig. 1. Diagrama general del supercomplejo fotosintético en bacterias púrpura
Las bacterias púrpura fotosintéticas son un grupo de Proteobacterias fotosintetizadoras anaerobias, las cuales utilizan como donadores de electrones succinatos, lactatos y eventualmente H2 o H2S a bajas concentraciones. Su aparato fotosintético, al igual que en los demás grupos biológicos, se encuentra embebido en invaginaciones de la membrana interna celular, las cuales generan un volumen confinado de periplasma donde se genera un gradiente de iones H+ requerido para la síntesis posterior de ATP.
El aparato fotosintético comprende un conjunto de proteinas transmembrana con funciones fínamente acopladas, e incluye complejos antena (LHII y LHI) destinados a la captura de energía lumínica y transporte de la misma al centro de reacción RC; el centro de reacción RC propiamente dicho, destinado a recibir y transferir la energía desde pigmentos fotosintéticos especiales hasta quinonas aceptoras a lo largo de una cadena de transporte de electrones, (Electron Transport Chain, ETC), incluye complejos cytocromo BC1, responsable de la generación PMF por medio de receptores especiales de quinonas; y finalmente complejos moleculares ATP synthase, que utilizan la PMF generada en la etapa anterior para ensamblar ATP.
El conjunto de subunidades conforman un supercomplejo transportador y almacenador de energía en forma de ATP y NADPH, moléculas fundamentales tanto para la activación de las cadenas de síntesis biológica como en los procesos de crecimiento, reproducción celular y en general en todas las funciones bioquímicas y fisiológicas demandantes de energía.
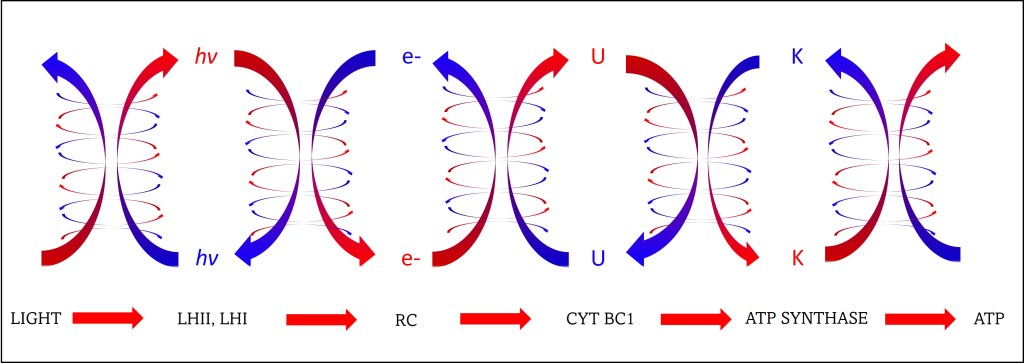
Fig. 2. Transferencia de energía entre complejos fotosintéticos. En la figura, “U” y “K” representan energías potencial y cinética, respectivamente.
CAPTURA DE ENERGÍA: COMPLEJOS ANTENA LHII Y LHI
La energía lumínica es capturada en arreglos concéntricos de carotenos y bacterioclorofilas (BChl) , localizados en complejos proteínicos octaméricos transmembrana, denominados complejos antena LHII (Light Harvest complex II). La energía es enrutada al interior de LHII de 500 a 800 a 850 nm para posteriormente exitar arreglos igualmente concéntricos de BChl ´s en LH1 a 875 nm.

Fig. 3. Light harvesting Complex II en Rhodoblastus acidophilus
Fuente: Photosynthesis | The Purple Photosynthetic Bacterial Light Harvesting System. Richard J. Cogdell, Tu C. Nguyen-Phan, in Encyclopedia of Biological Chemistry (Third Edition), 2021
Note en la figura la disposición en empalizada de los carotenos (moléculas café), y las disposiciones en anillos concéntricos de las 8 BChl´s paralelas a la superficie de la membrana plasmática (BChl´s violetas) y de las 16 perpendiculares a la misma (BChl´s azules).
La transferencia de energía entre y dentro de anillos se realiza mediante “RESONANCIA DE FÖRSTER”, FRET, un tipo particular de resonancia cuántica no radiativa, que requiere distancias muy reducidas entre moléculas donadora y aceptora, y overlapping al menos parcial de la frecuencia de emisión del donador y frecuencia de exitación del aceptor, ver figuras 4 y 5.
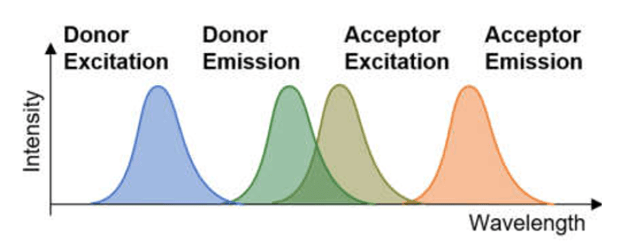
Fig. 4. Overlapping parcial entre frecuencia de emisión del donador y exitación del aceptor

Fig. 5. Dependencia de la distancia entre moléculas donadora y aceptora para los valores de transferencia de energía.
A diferencia de otros tipos de resonancia (Resonancia de Dexter), la Resonancia de Förster no mueve electrones exitados de donador a aceptor ni modifica su spin cuántico, por lo cual se define como una resonancia Singlet-Singlet no radiativa.
Partiendo de una exitación de un electrón de HOMO a LUMO en la molécula donadora, ver fig. 6., se transfiere posteriormente su energía a moléculas adyacentes mediante transferencia no radiativa.
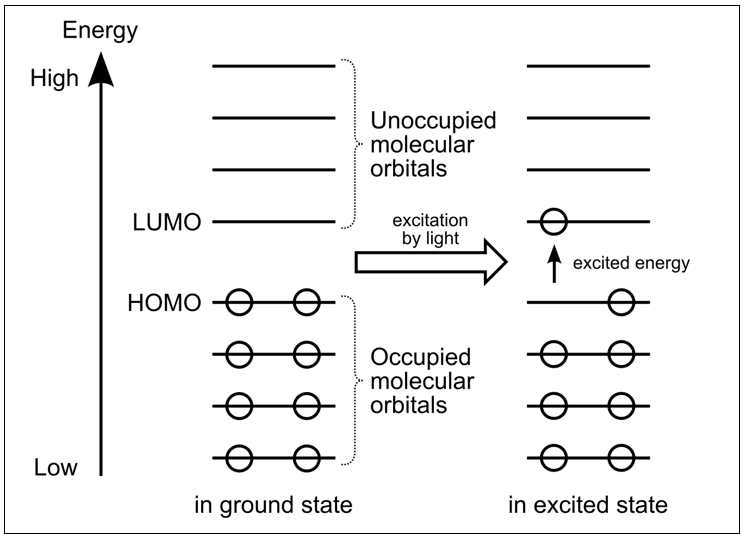
Fig. 6. Exitación energética de HOMO a LUMO
En este caso, la molécula donadora retorna un electrón de LUMO (Lowest Unoccupied Molecular orbital) a Homo (Highest Occupied Molecular Orbital) mientra que en la molécula aceptora uno de sus electrones eleva su energía de HOMO a LUMO.
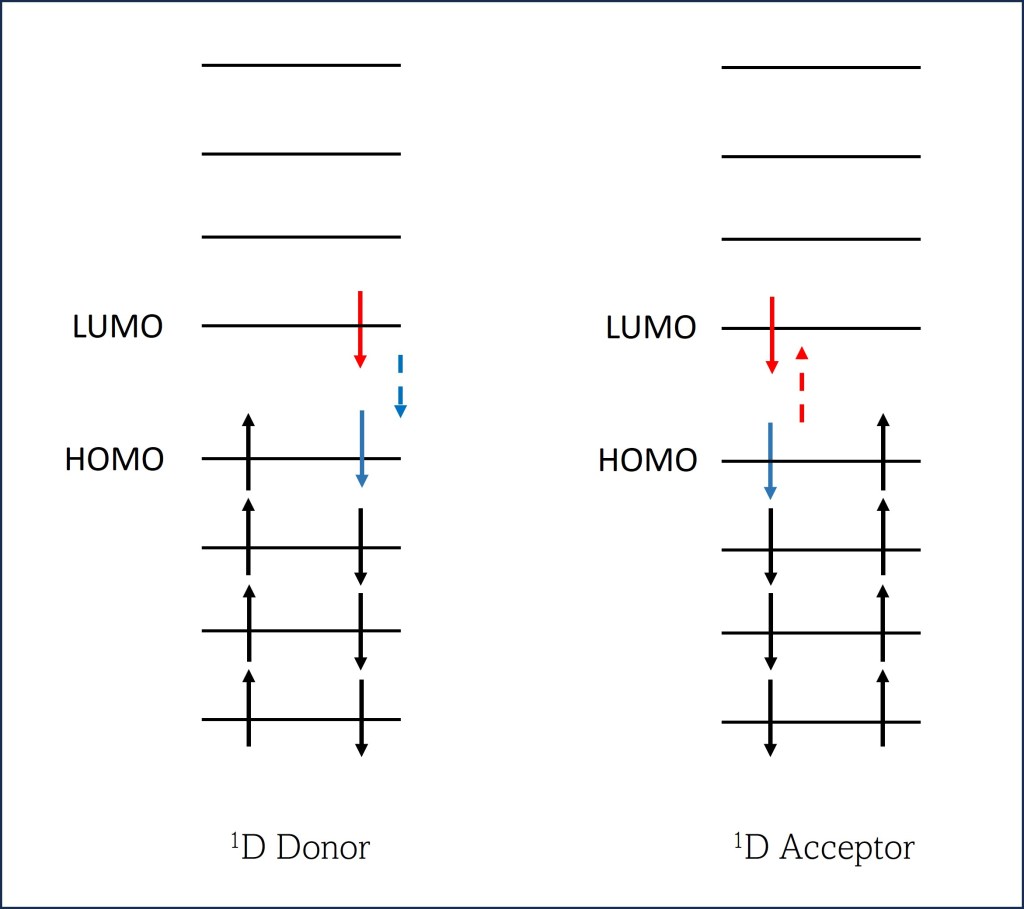
Fig. 7. Singlet-Singlet Förster Resonance Energy Transfer
Al menos dos propiedades cuánticas adicionales y probablemente una tercera son aprovechadas por los Complejos Antena para ejecutar su función: Superposición Cuántica, Coherencia Cuántica y eventualmente Superradiancia Cuántica.

Fig. 8.Transferencia de energía dentro de complejos (líneas azules) y entre complejos (Líneas rojas)
La Superposición Cuántica permite que multiples rutas probables coexistan tanto al interior de los complejos antena (flechas azules), como entre complejos (flechas rojas), garantizando que la energía sea transferida exitosamente y de manera rápida desde la molécula receptora del fotón hasta al Centro de Reacción RC sin que en el tránsito se “pierda”en rutas erráticas. Una vez alcanzado RC, la energía es “leída” por el par especial de clorofilas y en este momento la función de onda colapsa o elimina las demás rutas probables.
La unidireccional en cada paso también se garantiza por la disminución de frecuencias = Disminución de energía = aumento de longitud de onda durante el tránsito en el complejo de antenas (ver fig. 4).
Sin embargo también se debe notar que la exitación de LHI se dá a una mayor frecuencia (865 nm) que la frecuencia del donador (875 nm).
Es posible que la Coherencia y la Superradiancia Cuántica actuando en conjunto sean el elemento responsable del fenómeno.

Fig. 9.Superradiancia Cuántica
En el caso de los complejos antena, es probable que las moléculas individuales emisoras respondan bajo coherencia cuántica de forma espontánea , actuando cooperativamente como una única entidad. La coherencia lleva a que el grupo emita luz en un único haz coherente de alta intensidad. Este haz adquiere una intensidad N veces más fuerte que la intensidad esperada de un grupo de partículas independientes. En la figura 9 la variable N corresponde al número de partículas coherentes. Este efecto NO es basado en NINGUNA interacción entre las moléculas implicadas; es el resultado de las propiedades SIMÉTRICAS de la interacción del conjunto de partículas con el campo electromagnético lumínico global.
En resumen, los complejos antena, como parte del supercomplejo fotosintético, garantizan que la energía incidente de un fotón sea transmitida de manera eficiente hasta el centro de reacción RC, donde un par especial de clorofilas se exitarán al punto de liberar un electrón iniciando una cadena de transporte de electrones (ETC) que al final del proceso se acumulará en moléculas especializadas. El tránsito de fotónica a electrónica se ejecuta en el centro de reacción RC.
Orlando Rodríguez
Julio2, de 2025
LECTURAS RECOMENDADAS
Hu X, Schulten K. Model for the light-harvesting complex I (B875) of Rhodobacter sphaeroides. Biophys J. 1998 Aug;75(2):683-94. doi: 10.1016/S0006-3495(98)77558-7. PMID: 9675170; PMCID: PMC1299743.
Hu X, Ritz T, Damjanović A, Autenrieth F, Schulten K. Photosynthetic apparatus of purple bacteria. Q Rev Biophys. 2002 Feb;35(1):1-62. doi: 10.1017/s0033583501003754. PMID: 11997980.
Miroslav Z. Papiz, Anna M. Hawthornthwaite-Lawless, Steve M. Prince, Gerry McDermott, Andy A. Freer, Neil W. Isaacs, Richard J. Cogdell,
A model for the photosynthetic apparatus of purple bacteria,
Trends in Plant Science,
Volume 1, Issue 6,
1996,
Pages 198-206,
ISSN 1360-1385
